Презентація на тему:
Міграція
Завантажити презентацію





 складаються із актину, який може існувать в...")









 ек...")





Міграція
Завантажити презентаціюПрезентація по слайдам:

Процеси клітинного морфогенезу, міжклітинна комунікація та міграція клітин в нормі та за патологічних станів

До процесів клітинного морфогенезу відносяться: - - зміни клітин, зв’язані з переміщенням клітин на підложці (in vitro) процеси ендоцитозу, процеси утворення контакту клітин з підложкою та іншими клітинами, процеси орієнтування та міграції клітин. В ході морфогенетичних реакцій змінюються рецептори на поверхні мембрани (в тому числі і рецепторні адгезивні молекули).

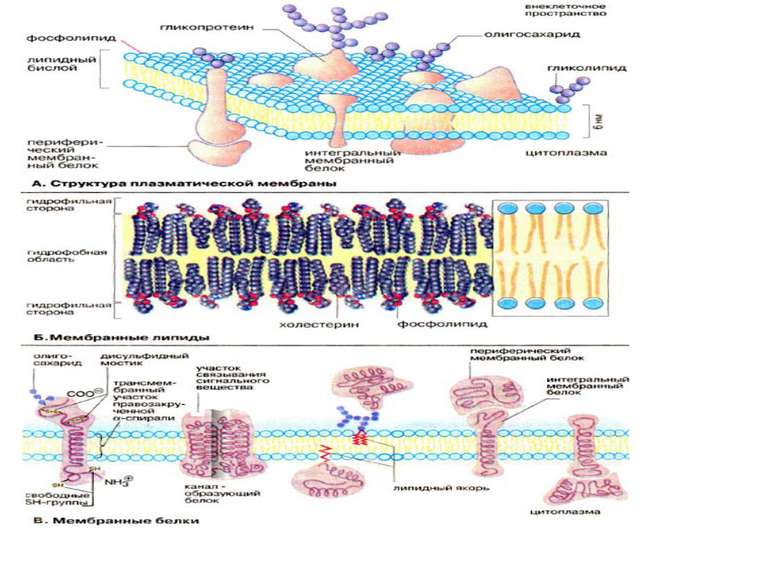
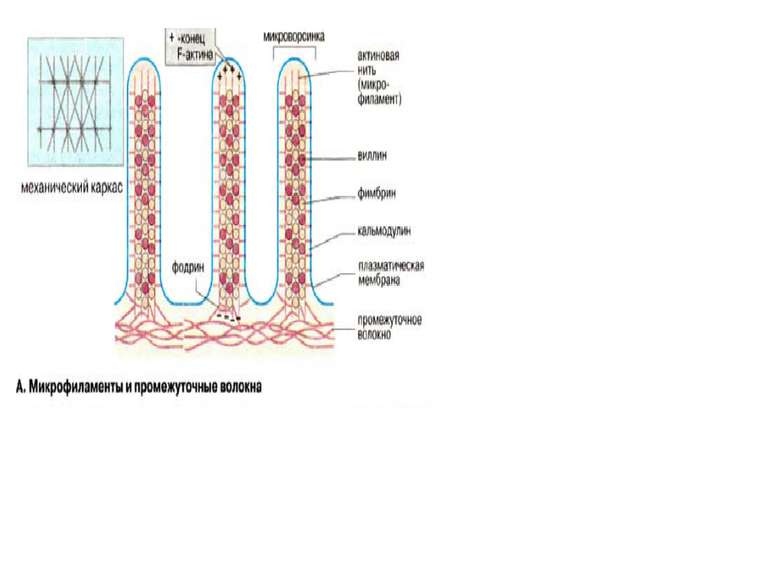
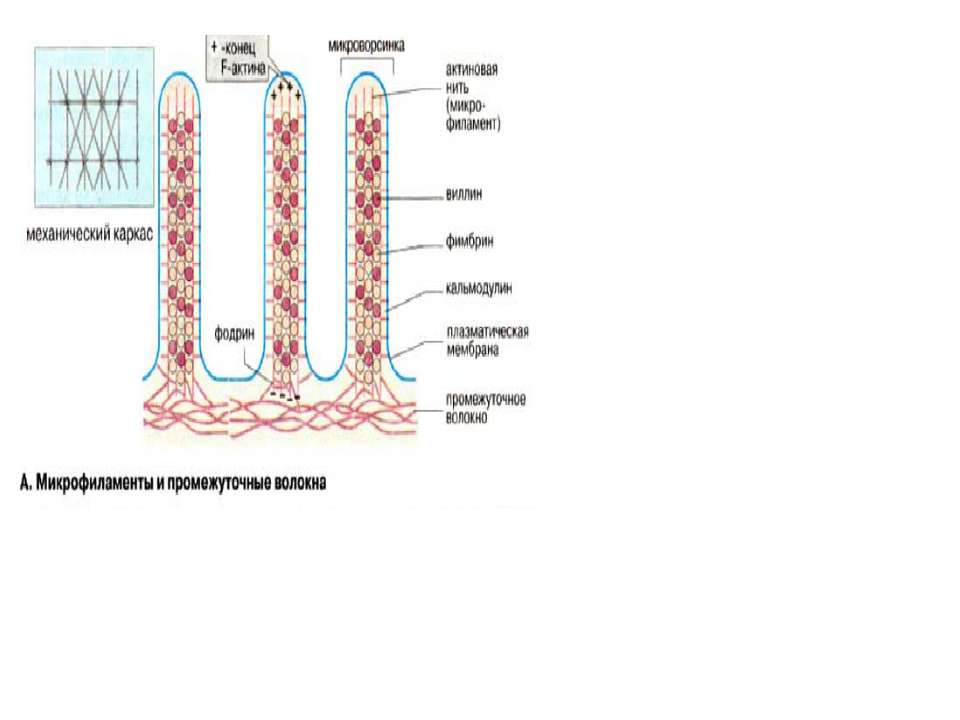
Клітинна поверхня розміщена на межі клітини зі зовнішнім середовищем і містить в собі дві основні структури: клітинну мембрану і підмембранний кортикальний шар. Кортикальний шар – це зона під клітинною мембраною, де є фібрили, але відсутні рибосоми і везикулярні компоненти. В клітинах вирізняють три групи фібрилярних структур: мікрофіламенти (4-6 нм), проміжні філаменти (інтермедіальні-7-12 нм) і мікротрубочки (25нм). Мікрофіламенти містять актин – основний білок клітин нем’язевого походження. Актин буває поляризований і неполяризований. Структури, які утворюються актином та іншими білками (віментін, філамін) здатні скорочуватись. Структура кортикального шару клітин в культурі може дуже швидко змінюватись в результаті поляризації і деполяризації мікрофіламентів, за рахунок чого клітини надзвичайно динамічні.

Плазматична мембрана являє собою біслой ліпідів з вбудованими білками (периферичні та інтегральні) Білки клітинної мембрани класифікують за функціональною приналежністю: Структурні білки: - надають клітині та органелам певну форму - надають мембрані та плазмолемі ту або іншу механічну функцію (еластичність, збудливість) забезпечують зв’язок мембрани з цитоскелетом, або у випадку ядерної мембрани з хромосомами
 складаються із актину, який може існувать в...")
Мікрофіламенти (актинові волокна) складаються із актину, який може існувать в вигляді мономера (G-актин, «глобулярний актин») або полімера (F-актин, «фібрилярний актин»). G-актин — асимметричний глобулярний білок (42 кДа), який складається з двох доменів. По мірі підвищення іонної сили G-актин зворотньо агрегує, утворюючи скручений в спіраль полімер, F-актин. Молекула G-актина несе зв’язану молекулу АТФ, яка при переході в F-актин, повільно гідролізуєтся до АДФ, т.е. F-актин проявляє властивості АТФ-ази.При полімеризації G-актина в F-актин орієнтація всіх мономерів однакова, тому F-актину властива полярність

Актинасоційовані білки. В цитоплазмі клітин є більше 50 різних типів білків, які специфічно взаємодіють з G-актином та F-актином. Ці білки виконують різні функції: регулюють об’єм G-актинового пулу (профілін), впливають на швидкість полімеризації G-актина (вілін), стабілізують волокна F-актина (фрагин, β-актинин), зшивають філаменти з іншими компонентами ( як наприклад, віліин, α-актинин, спектрин, MARCKS) розрушують подвійну спіраль F-актина (гельзолін). Активність цих білків регулюється іонами Са2+ та протеїнкіназами.

Тубулін Мікротрубочки побудовані із глобулярного білка тубуліна, який є димером α- та β-субодиниць (53 та55 кДа). α, β- Гетеродимери утворюють лінійні цепочки, які називаються протофіламентами. 13 протофіламентів утворюють циклічний комплекс. потім кільця полімеризуються в довгу трубку. Як і мікрофіламенти, мікротрубочки представляють собою динамічні полярні структури з (+)- та (-)-зарядженими закінченнями. (-)-кінець стабілізований за рахунок зв’язування з центросомою (центр організації мікротрубочок), в той час як для (+)-кінця характерна динамічна нестабільність.Він може або повільно рости, або швидко скорочуватись. Тубулінові мономери зв’язують ГТФ (GTP), який повільно гідролізується до ГДФ (GTP). З мікротрубочками асоційовані два види білків: структурні білки (MAP от англ. microtubuls-associated proteins) та білки-транслокатори.

Б. Білки інтермедіальних філаментів Структурними елементами проміжних волокон є білки, що належать до 5 подібних родин та мають виражену клітинну специфічність. Типовими представниками цих білків є цитокератини, десмін, віментін, кислий фібрилярний глиапротеїн [КФГП (GFAP)] та нейрофіламент. Вс і ці білки в центральній частині мають базову структуру, яка має назву эти белки имеют в центральной части базовую стержневую структуру, которая носит суперспіралізованої α-спіралі. димери асоціюють антипаралельно, утворюючи тетрамер. Агрегація тетрамерів по принципу "голова до голови" дає протофіламент. Вісім протофіламентів утворюють проміжне волокно. Полімеризація інтермедіальних філаментів, на відміну від мікротрубочок та мікрофіламентів приводить до утворення стійких неполяризованих структур Проміжні філаменти входять в склад різних клітин і утворюють 5 основних груп: кератинові (тонофіламенти) виявляються в епітеліальних клітинах різних типів, десмінові – в м’язевих клітинах, віментинові, в клітинах мезенхімального походження, нейрофіламенти, виявляються в нейронах, гліальні філаменти в клітинах глії.

Організація мікротрубочок Цитоскелет виконує три головні функції 1. Слугує механічним каркасом клітини, який надає клітині певну форму та забезпечує зв’язок між мембраною та органелами. Каркас є динамічною структурою, яка постійно поновлюється та реагує на подразники мікрооточення. 2. Діє як «двигун» для руху клітин. Компоненти цитоскелету визначають напрямок та кординацію руху поділу, змінуформи клітин в процесі росту, переміщення органел, рух цитоплазми 3. Слугує в якості «рельсів» для транспорту органел та інших крупних комплексів внутрі клітини.
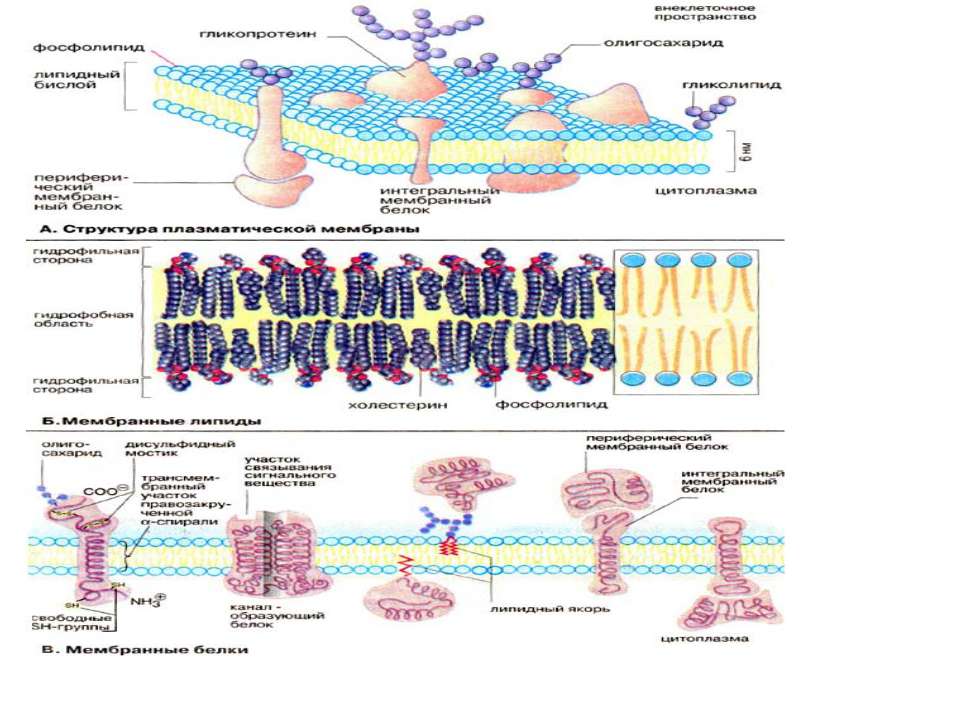

Всі біомембрани побудовані однаково; вони складаються із двох шарів ліпідних молекул біля 6 нм, в які вбудовані білки. Деякі мембрани містять, вуглеводи, зв’язані з ліпідами та білками. Співвідношення ліпіди : білки : вуглеводи є характерним для клітин або мембрани та суттєво варіює в залежності від типу клітин або мембран. Компоненти мембран утримуються нековалентними зв’язками, за рахунок чого вони мають властивість лише відносної рухливості тобто можуть дифундувать в межах ліпідного бішару. «Текучесть» мембран залежить від ліпідного складу та температури мікрооточення. Із збільшенням вмісту ненасичених жирних кислот «текучесть» збільшується, так як наявність подвійних зв’язків сприяє порушенню напівкристалічних мембранних структур. Рухливими є і мембранні білки. Якщо білки не закріплені в мембрані, вони «плавають» в ліпідному бішарі як в рідині, і тому існує таке поняття як рідинно-мозаїчна структура біомембрани. В той час як «дрейф» в площині мембрани відбувається достатньо легко, перехід білків із зовнішньої сторони мембрани на внутрішню («фліп-флоп») неможливий, а перехід ліпідів відбувається надзвичайно рідко. Для «перескоку» ліпідів необхідні спеціальні білки -транслокатори. Виключенням є холестерин, який може легко переходить з однієї сторони мембрани на іншу.

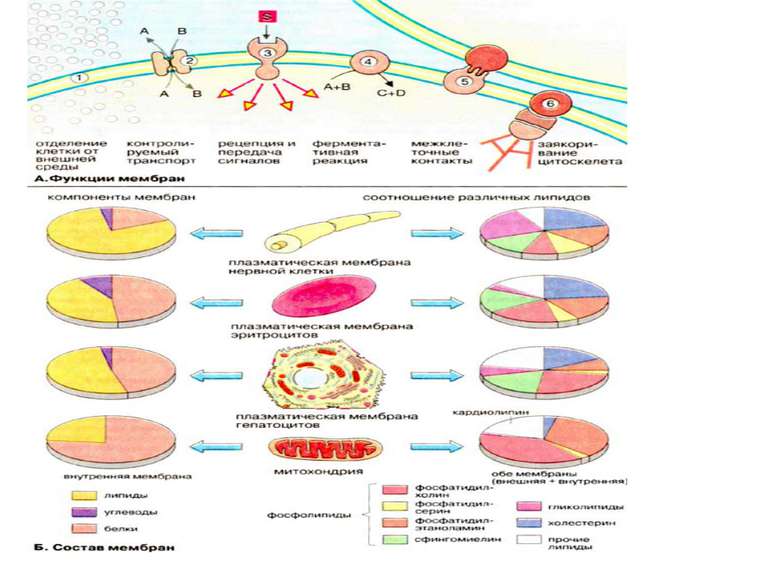
Мембранні ліпіди В мембранах містяться ліпіди трьох класів: фосфоліпіди, холестерин та гліколіпіди. Найбільш важлива группа, фосфоліпіди, включає фосфатітілхолін (лецитин), фосфатідилетаноламін, фосфатідилсерин, фосфатідилінозит та сфінгомієлин. Холестерин присутній у внутрішньоклітинних мембранах клітин (за виключенням внутрішньої мембрани мітохондрій). Гліколіпіди входять в склад багатьх мембран (наприклад, в зовнішній шар плазматичних мембран). В склад гліколіпідів входять вуглеводні функціональні групи, які орієнтуються в водну фазу. Ліпіди мембран являють собою амфіфільні молекули з полярною гідрофільною головкою та неполярним ліпофільним хвостом. В водному середовищі вони агрегують за рахунок гідрофобних взаємодій та вандерваальсових сил.

Мембранні білки Протеїни можуть зв’язуватись з мембраною різним шляхом. Інтегральні мембранні білки мають трансмембранні спіралізовані домени, які однократно або багатократно пересікають ліпідний бішар. Такі білки «прочно» зв’язані з ліпідним оточенням. Периферийні мембранні білки утримуються на мембрані за допомогою ліпідного «якоря» та зв’язані з іншими компонентами мембрани; найчастіше ассоційовані з інтегральними мембранними білками. У інтегральних мембранних білків фрагмент пептидной цепочки, який пересікає ліпідний бішар, складається із 21-25 переважно гідрофобних амінокислот, які утворюють праву α-спіраль з 6 або 7 витками (трансмембранна спіраль). Периферичні легко відділяються від ліпідного шару при мінімальних обробках, інтегральні білки вбудовані і їх можна видалити лише при розрушенні мембрани. До деяких мембранних ліпідів та білків (гліколіпіди та глікопротеїни) приєднані олігосахаридні ланцюги. Плазматична мембрана може поповнювати свій склад (старі молекули можуть інтерналізуватись внутріклітинно та деградувати).

Класифікація мембранних білків може бути також проведена за їх функціональною приналежністю. Структурні білки: надають клітині та органелам певну форму надають мембрані та плазмолемі ту або іншу механічну функцію (еластичність, збудливість) забезпечують зв’язок мембрани з цитоскелетом, або у випадку ядерної мембрани з хромосомами Транспортні білки Проникність мембрани обумовлена ліпідним бішаром, хоча його можливості обмежені невеликою групою молекул, а основна група молекул переміщається через мембрану через транспортні білки, які поділяються на односторонні та двохсторонні. Створюються стійкі транспортні потоки певних речовин через мембрану, а також транспорт іонів призводить до змін трансмембанного потенціалу Мембранні рецептори - інтегральні компоненти мембрани, здатні активуватись зв’язуючись з специфічними лігандами. рецептори лектинів, які зв’язують певні моносахариди (наприклад конковалін А) рецептори кінцевих груп олігосахаридів рецептори сенсебілізованих лімфоїдних клітин, які зв’язують відповідні антигени (імуноглобулінові молекули) рецептори білкових гормонів та нейромедіаторів (інсулін) рецептори бактерільних токсинів (холерний токсин) адгезивні рецептори (інтегрини, селектини, кадгерини, імуноглодулінподібні молекули)
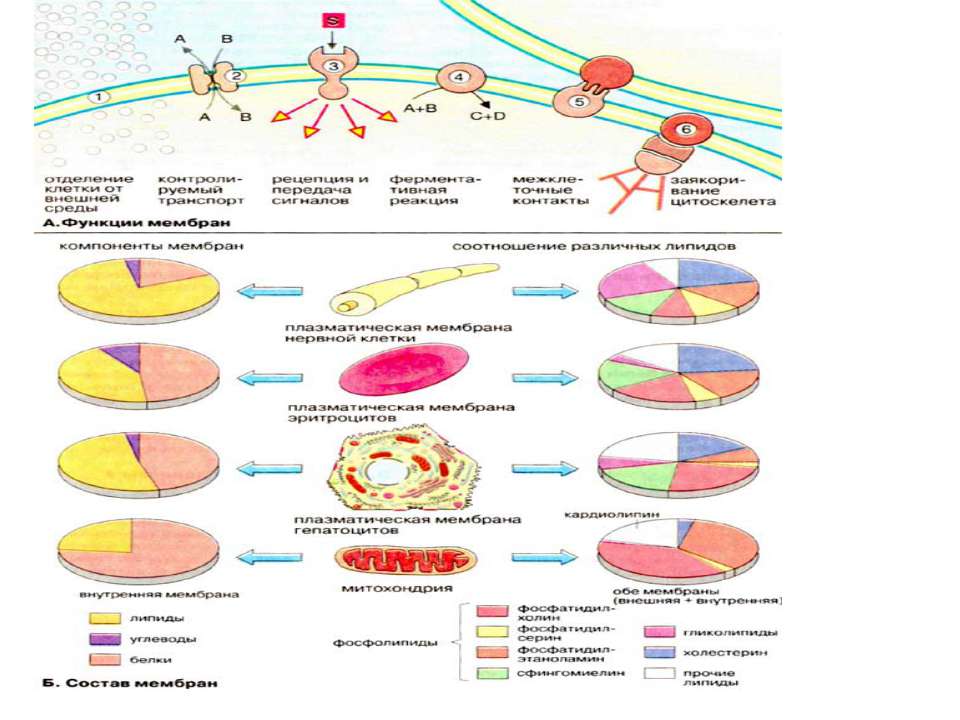

Функції біомембран Біомембрани виконують слідуючі функції: 1. Обмеження та обособлення клітин та органел. Обособлення клітин від міжклітинного середовища забезпечується плазматичною мембраною, яка захищає клітину від механічного та хімічного подразнення. Плазматична мембрана забезпечує також збереження різниці концентрацій метаболітів та неорганічних іонів між внутріклітинним та зовнішнім середовищем. 2. Контрольований транспорт метаболітів та іонів визначає внутрішнє середовище, що є суттєвим для гомеостазу, тобто підтримання постійної концентрації метаболітів та неорганічних іонів, а також інших фізіологічних параметрів. Регульваний селективний транспорт метаболітів та неорганічних іонів через пори та опосередковано білками-переносниками стає можливим із-за обособлення клітин та органел за допомогою мембранних систем. 3. Сприйняття позаклітинних сигналів, їх внутріклітинна передача та ініціація сигналів. 4. Ферментативний каталіз. В мембранах на межі між ліпідною та водною фазами локалізовані ферменти. Якраз тут відбуваються реакції із неполярними субстратами Прикладом може слугувати біосинтез ліпідів та метаболізм неполярних ксенобіотиків. В мембранах локалізовані найбільш важливі реакції енергетичного обміну, такі, як окислювальне фосфорилювання та фотосинтез (рослинні клітини) 5. Контактна взаємодія з міжклітинним матриксом та взаємодія з іншими клітинами при при злитті клітин та утворенні тканин. 6. Заякорення цитоскелету, яке забезпечує підтримання форми клітин та органел та клітинної рухливості.
 ек...")
Взаємодія між клітинами в організмі відбувається за трьома механізмами: а) екстраклітинна взаємодія - опосередковано гормонами, ростовими факторами, нейротрансміттери та цитокіни, б) внутріклітинна взаємодія через іншу групу посередників (Са, рН, кераміди, оксид азоту,с-АМР, реактивний кисень), активує системи сигнальної трансдукції і модулює в)міжклітинну взаємодію через gap junction channels (3), яка відбувається через субклас міжклітинних комунікаційних молекул.

Слід підкреслити, що в фізіологічних умовах ліпіди, які утворюють мембрану, знаходяться переважно в рідко-кристалічному виді. Це означає, що кожна молекула внутрі свого моношару має високу рухливість, але перехід молекул із одного ліпідного моношару в інший спостерігається дуже рідко. Зміни мікров’язкості ліпідного шару можуть бути результатом зміни співвідношення різних хімічних класів ліпідів в мембрані. Багато рецепторів здатні рухатись в ліпідному шарі, проте існують механізми, які обмежують цю рухливість: - заякорення рецепторів ззовні, тобто прикріплення їх до якоїсь структури ззовні від мембрани, заякорення рецепторів внутрі клітини до кортикальних структур, що розміщені під мембраною агрегація рецепторів в площині мембрани.

Свіжеотримані первинні клітини і висаджені в культуру мають сферичну форму. Механізми, які сприяють розпластання клітин на підложках слідуючі: Початкове прикріплення – клітина прикріплюється нижньою поверхнею до підложки, проте ніяких виростів ще нема, лише в області контакту мембрана потовщена, але кортикальний шар ще не змінений. Утворення ламелярної цитоплазми – відсутні крупні органели і існують маса точок прикріплення до підложки. Стадія поляризації. Різні типи псевдоподій, які утворюються при розпластанні клітин на поверхні мають ряд загальних властивостей: -фокальні контакти між клітиною і підложкою утворююються на поверхні псевдоподій, але не на поверхні інших частин клітини -в цитоплазмі псевдоподій утворюється центробіжне натягнення, інертні частинки, які попадають на поверхню псевдоподій і ламелярної цитоплазми, прикріплююються до цієї поверхні і переміщуються по ній центробіжно -розвиток натягнення, в прикріпленій псевдоподії зберігається натягнення, бо при їх розрушенні псевдоподії та ламілярна цитоплазма моментально скорочується очистка від склеєних рецепторів, які після приєднання лігандів агрегують відбувається слідуючим чином: спершу очищуються кінці псевдоподій по центробіжному параметру (від кінця псевдоподій до центру).

механізм псевдоподіальних реакцій може включати в себе декілька стадій: - початковий етап- це іммобілізація группи рецепторів мембрани - утворення такої групи рецепторів може індукувати кортикальні зміни, що приводять до появи псевдоподій; -якщо утворена група імобілізованих рецепторів знаходиться на поверхні псевдоподій, то вони індукують іншу групу кортикальних змін, що приводять до заякорення цієї групи рецепторів внутрі і до їх з’єднання з актиновими мікрофіламентами, в результаті в місті контакту розвивається цетробіжний натяг в залежності від умов реакції прикріплення або натягу процес закінчується або прикріпленням і утворенням контакту, або центробіжним переміщенням імобілізованих рецепторів. Для трансформованих клітин характерне змінене формування мікровиростів та погіршене розпластання трансформованих клітин на поверхні. Ці характеристики можуть бути як причиною так і наслідком трансформованності клітин

Поляризовані трансформовані клітини також, як і нормальні здатні до активного переміщення по підложці, однак деякі статистичні характеристики цього переміщення можуть змінюватись при трансформації. Трансформовані клітини більш часто міняють напрямки при міграції (це показано мікрокінематографічними зйомками, а також по переміщенню клітин на підложці з колоїдним золотом). Такі зміни руху трансформованих клітин є вторинною ознакою дефектного прикріплення клітин до підложки: погано прикріплені вузькі активні місця легше відриваються від підложки, чим широкі добре прикріплені місця нормальних клітин. Стикання активного краю клітини, що рухається з краєм іншої клітини може призводити до трьох результатів: зупинці руху, підповзанню, або наповзанню клітин одна на іншу. При прямому спостереженні за поведінкою при русі трансформованих клітин було виявлено, що для трансформованих клітин характерне також пригнічення руху (зупинка, відсутність наповзання одна на іншу). Котнактне гальмування у трансформованих клітин частіше чим у нормальних супроводжується контактним паралічем та ретракцією. Ретракція у трансформованих клітин виражена більш сильніше, чим у нормальних і приводить до повного відриву від підложки та скороченню цитоплазматичних відростків, на яких розміщений активний край.

З точки зору псевдоподіальних реакцій різні культури трансформованих клітин можуть мати значне морфологічне різноманіття. Так, при дії на нетрансформовані клітин одним і тим же агентом можуть виникати різні трансформовані клони, які можуть мати різну морфологію, наприклад: -Одношарові або багатошарові культури, які складаються із погано розпластаних циліндричних клітин (зокрема фібробластів), в яких сусідні клітини паралельні одна іншій -Багатошарові культури, які складаються із погано розпластаних циліндричних полігональних або зірчатих клітин -Багатошарові клітини, які складаються в основному із округлих клітин -Одношарові культури, які складаються в основному із майже сферичних клітин, які сидять на підложці.
Схожі презентації











Категорії











