Презентація на тему:
11_10__3_Регуляція транскрипції еукаріот 11 10 11
Завантажити презентацію












 ділянки ...")












 - ферменти, що епігенетично модифікують г...")
 включає білки, що беруть участ...")

, Ybf2/Sas3, Sas2 і Tip60. Н.:...")
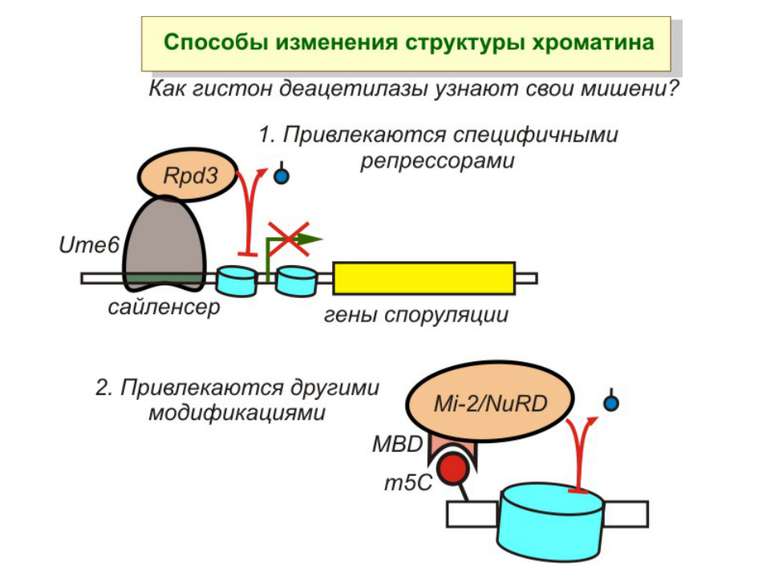
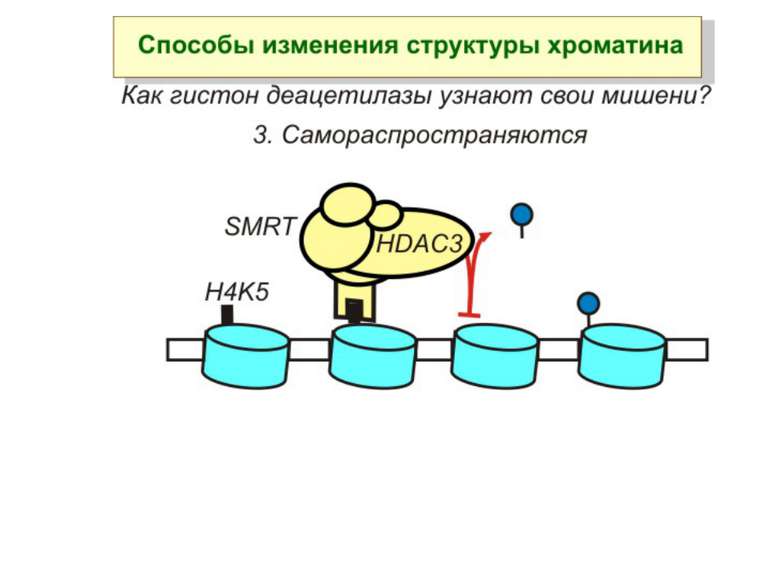

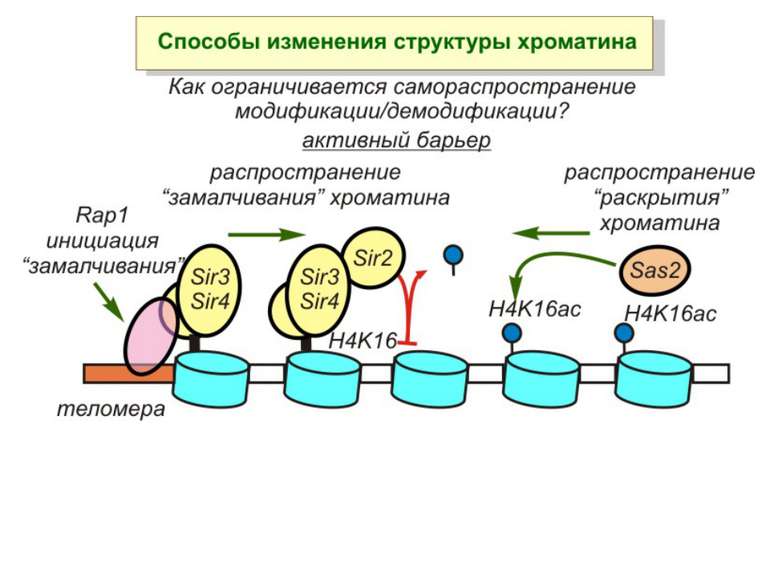

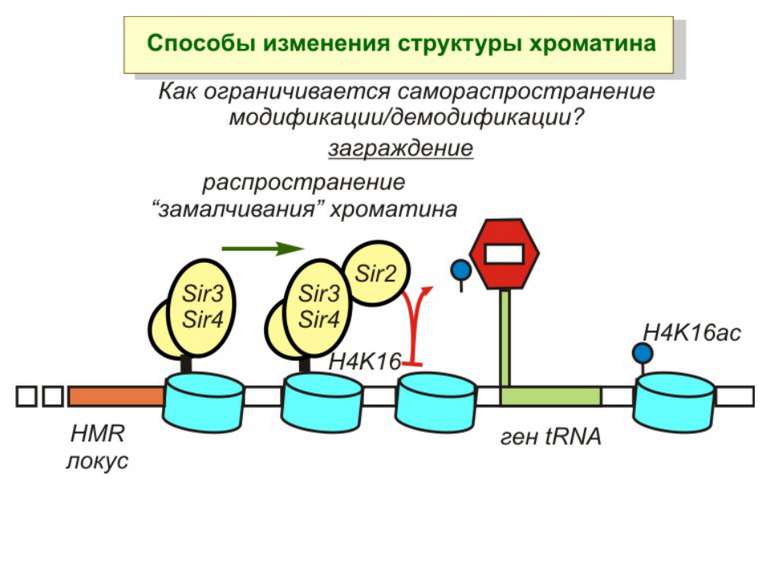





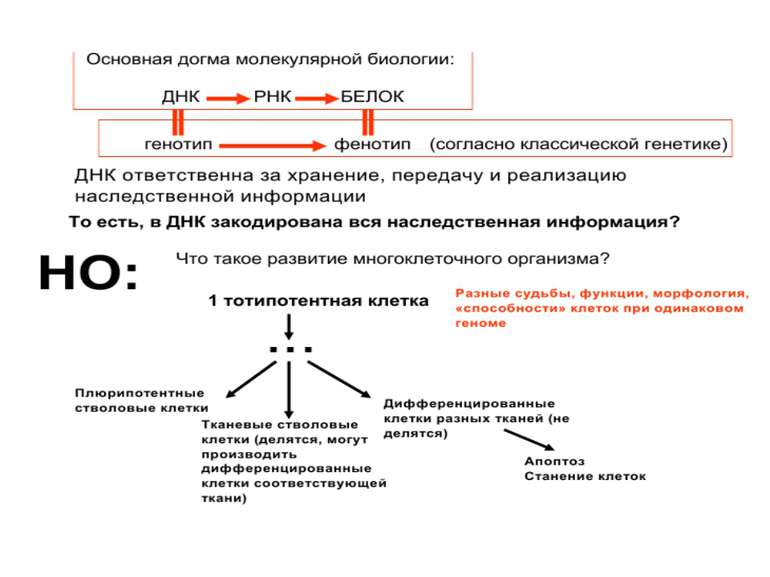
 визначається їх функцією")

























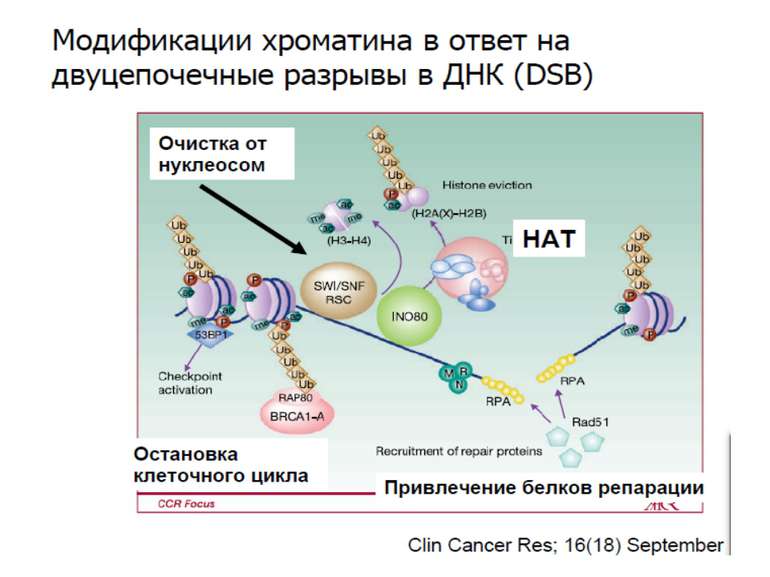




11_10__3_Регуляція транскрипції еукаріот 11 10 11
Завантажити презентаціюПрезентація по слайдам:

Регуляція транскрипції у еукаріот Роль епігенетичних механізмів
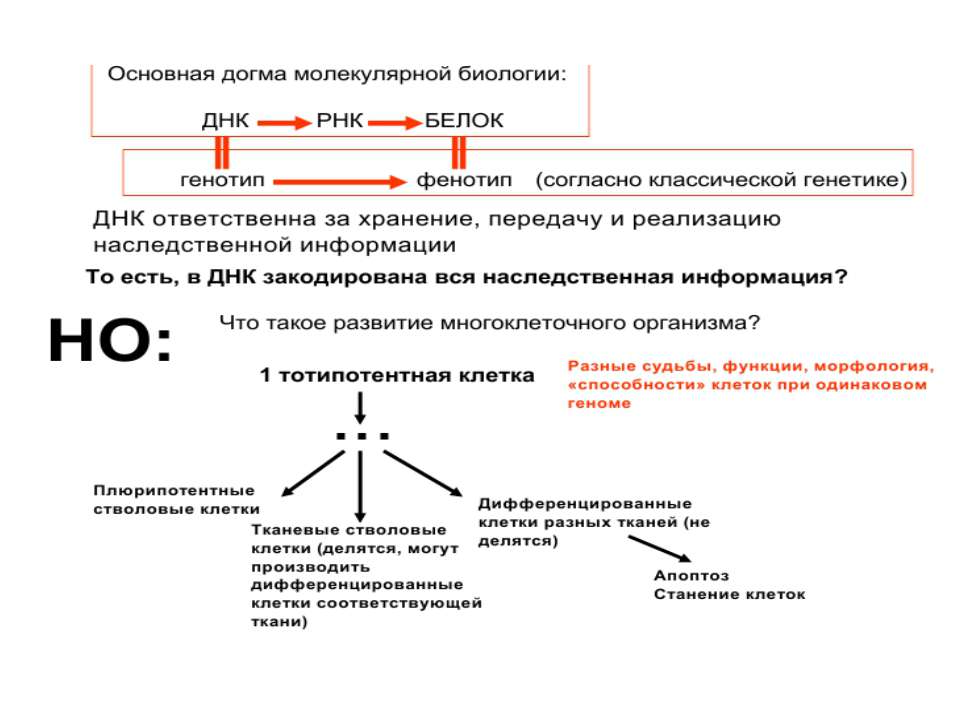

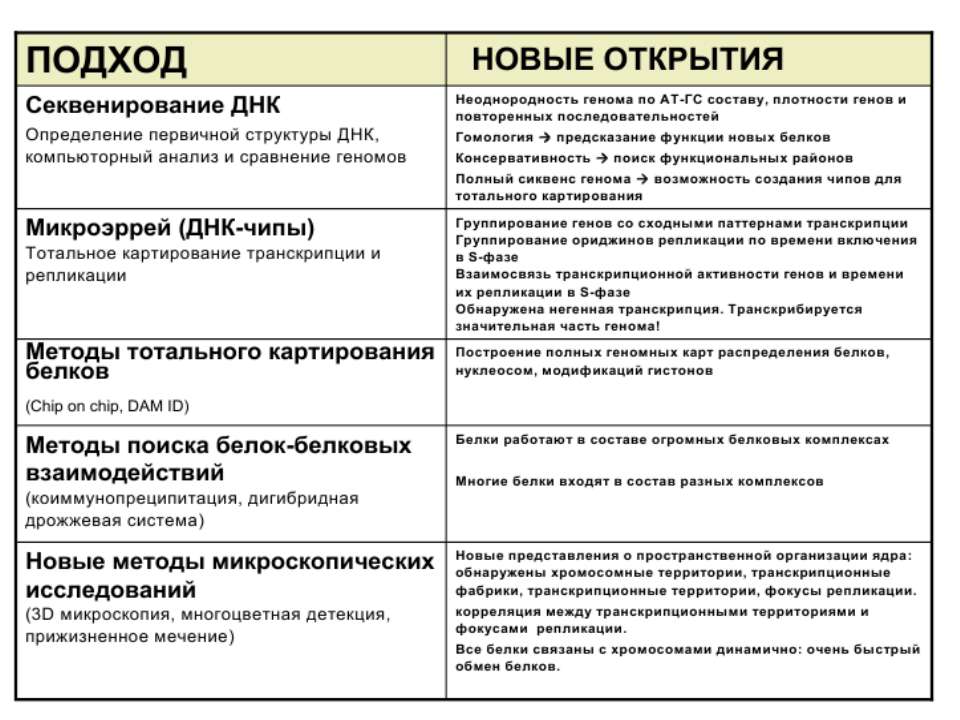



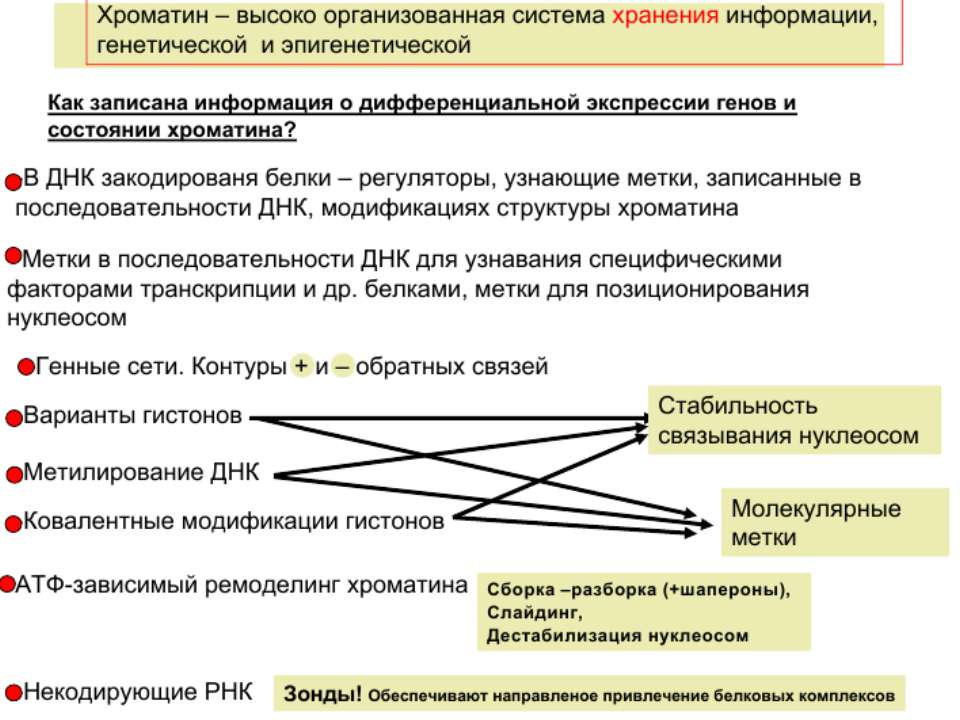
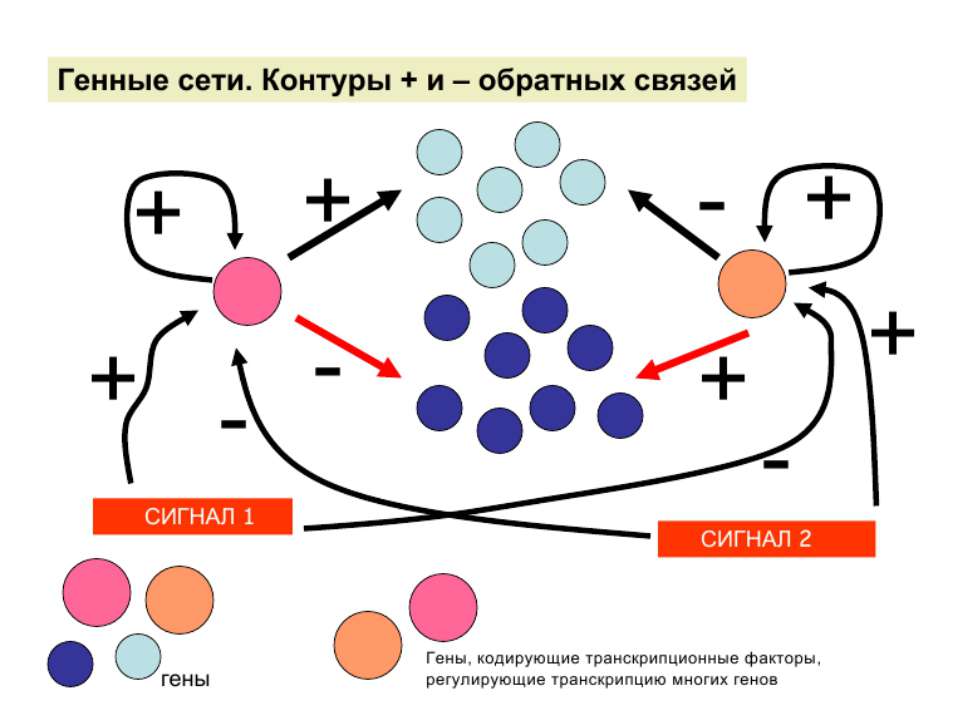
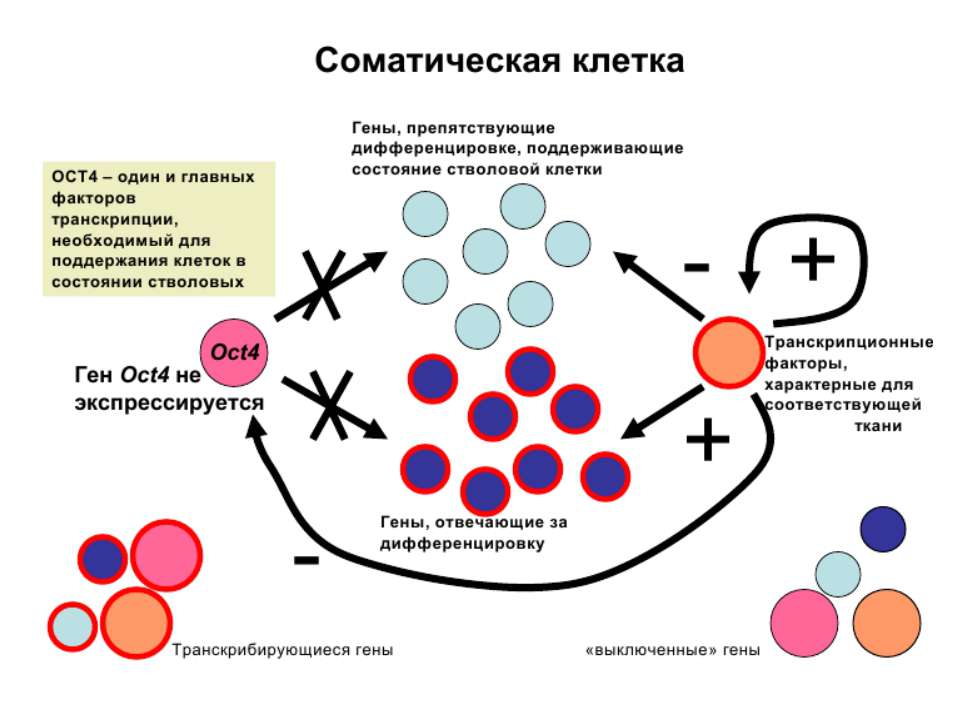
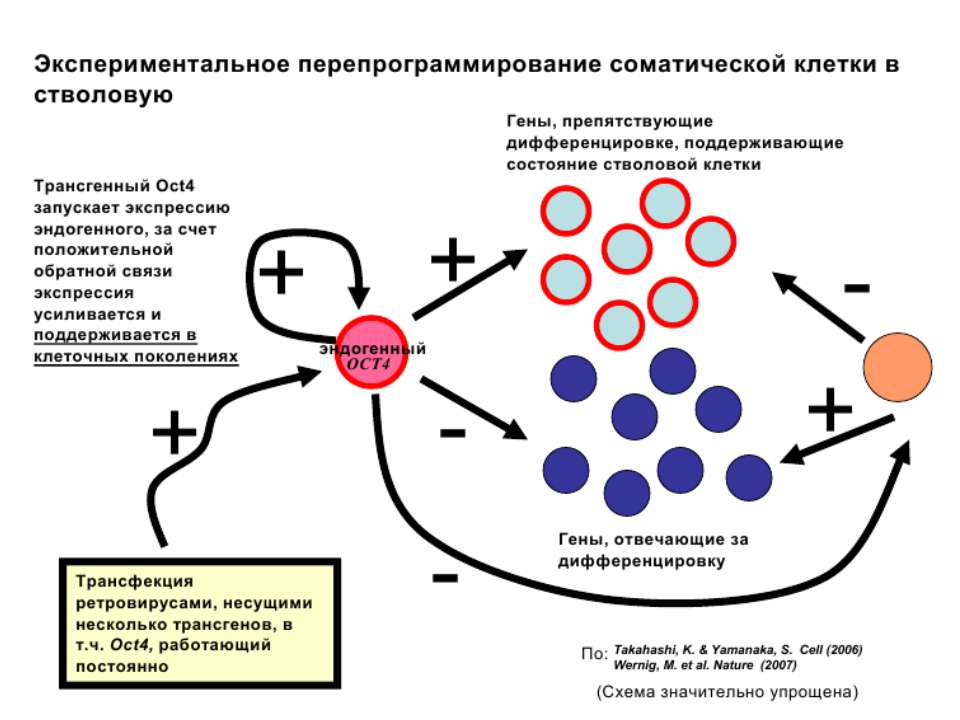
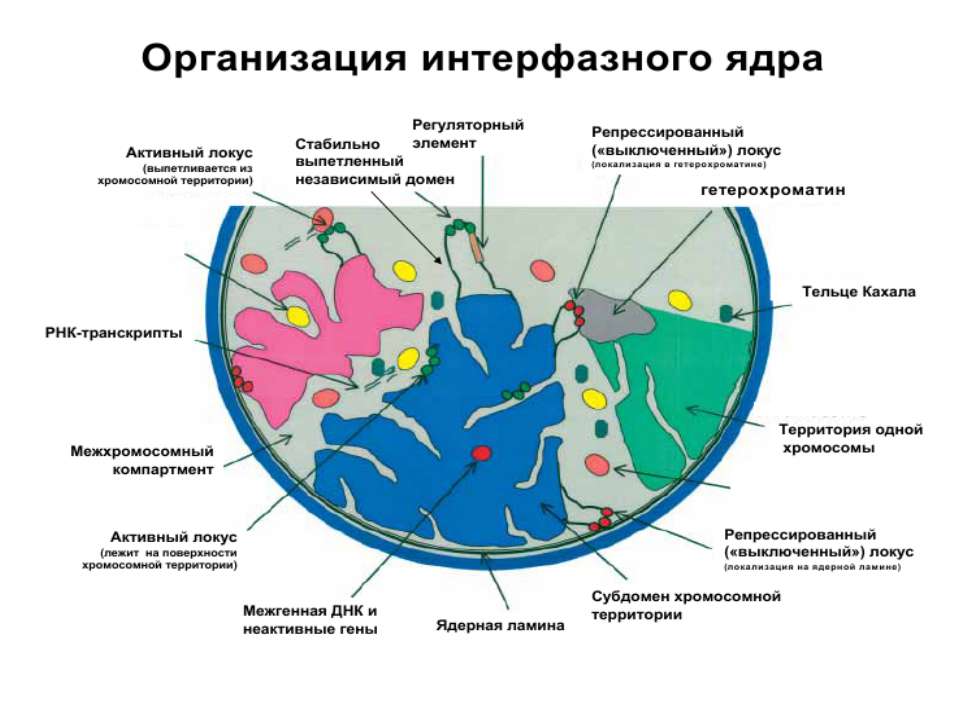
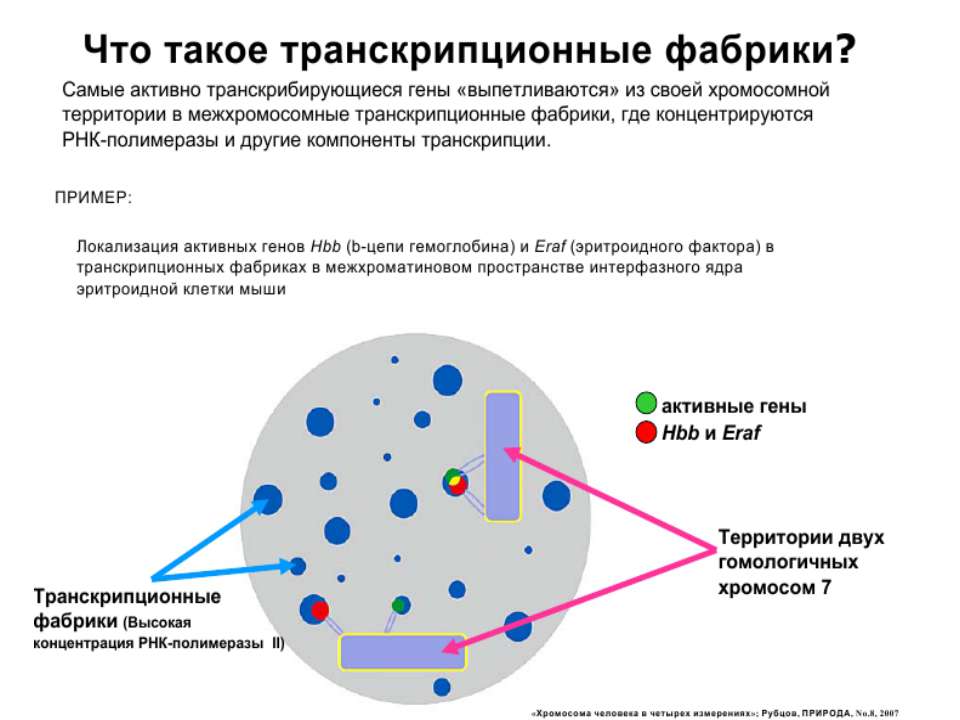

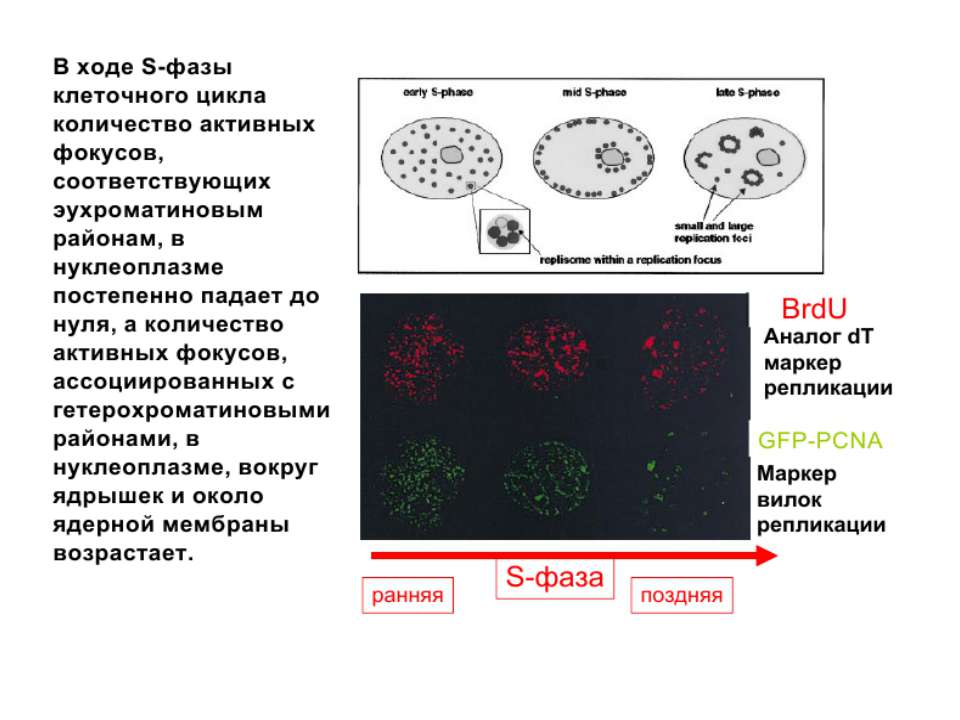


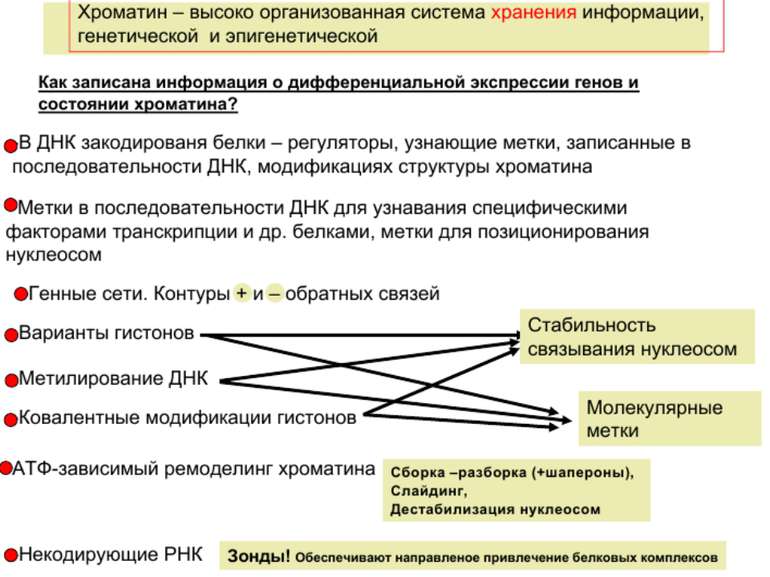
Регуляція експресії генів може здійснюватись на різних рівнях Хроматиновий рівень – конденсація хроматину та доменів хроматину Численні ділянки геному регулюються як єдині протяжні домени хроматину. На цьому рівні можлива епігенетична регуляція (епігенетичне успадкування стану експресії) Окремі гени У цій регуляцї беруть участь: Ковалентні модифікації ДНК Варіанти і модифікації гістонів Різні негістонові білки хроматину Просторова організація ядра Просторово-часова картина реплікації

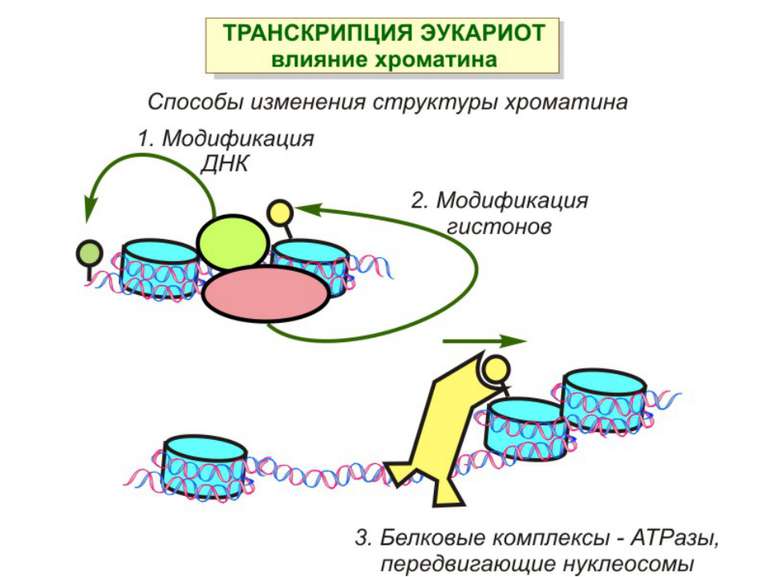
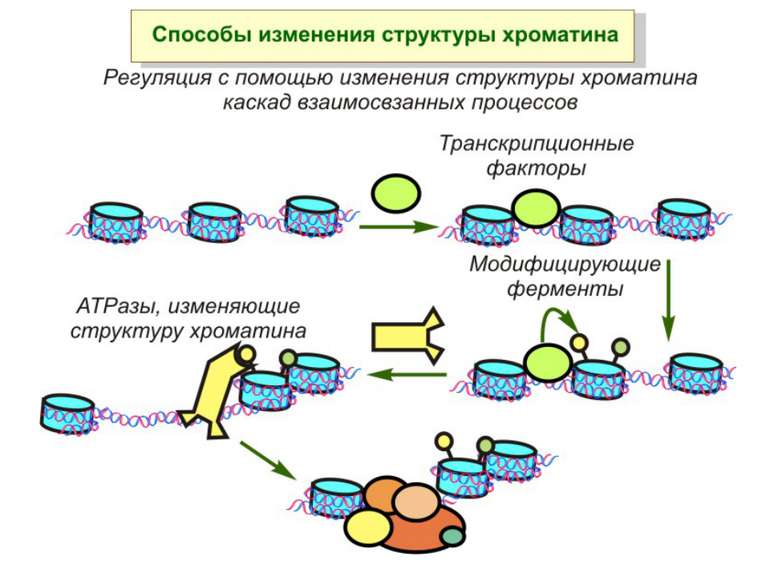
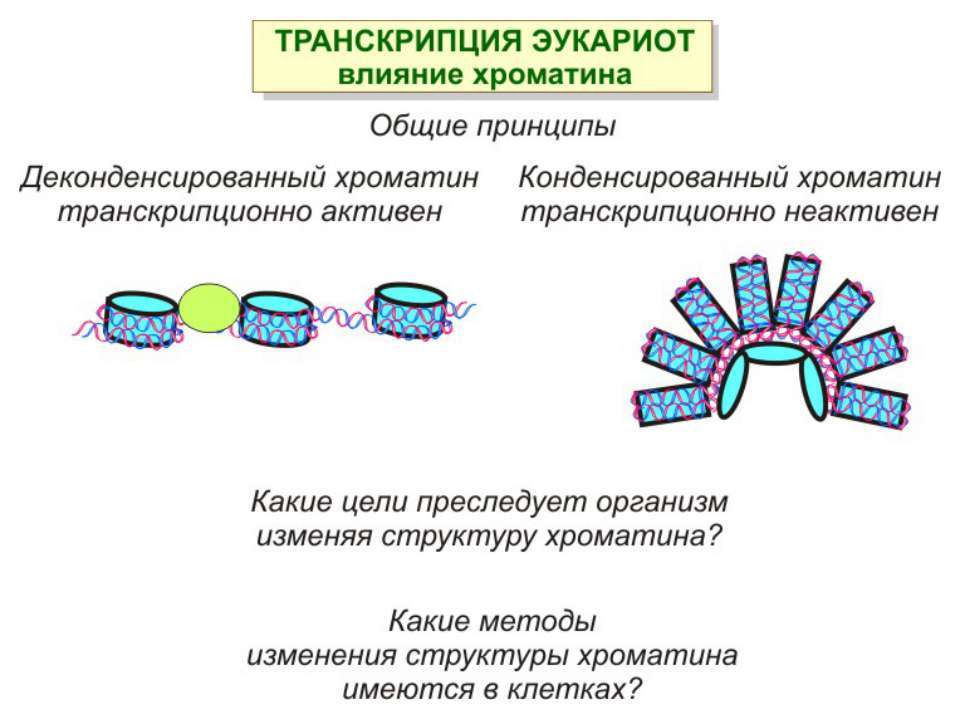
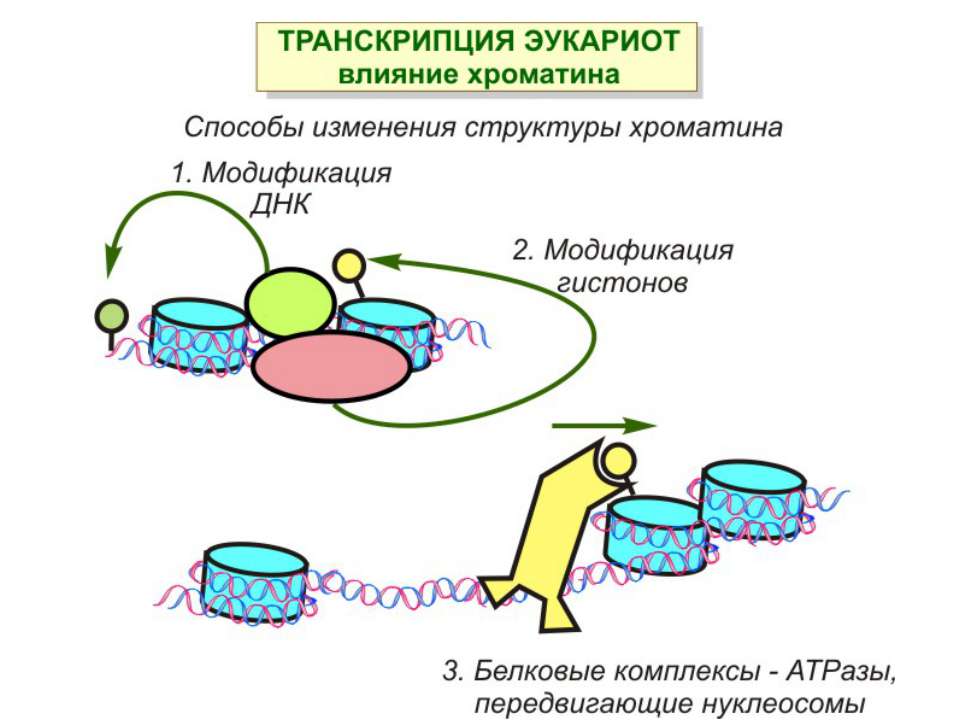
Механізми регуляції транскрипції еукаріот Зв’язування з ДНК регуляторних білків і факторів транскрипції. Модифікація ДНК (метилювання) і гістонів (ацетилювання, метилювання і ін.) Компактизація/декомпактизація хроматину, що здійснюється білковими комплексами

NB!!! Дотранскрипційний рівень !!!

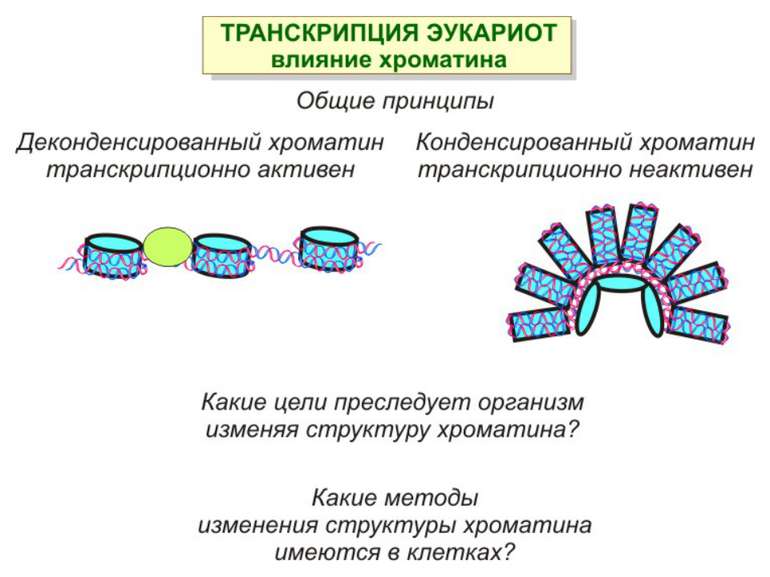
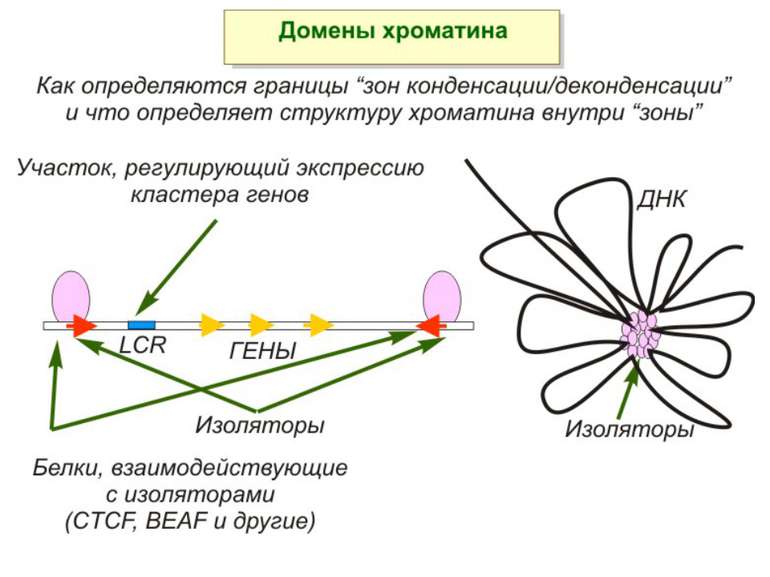
Хроматиновий рівень організації
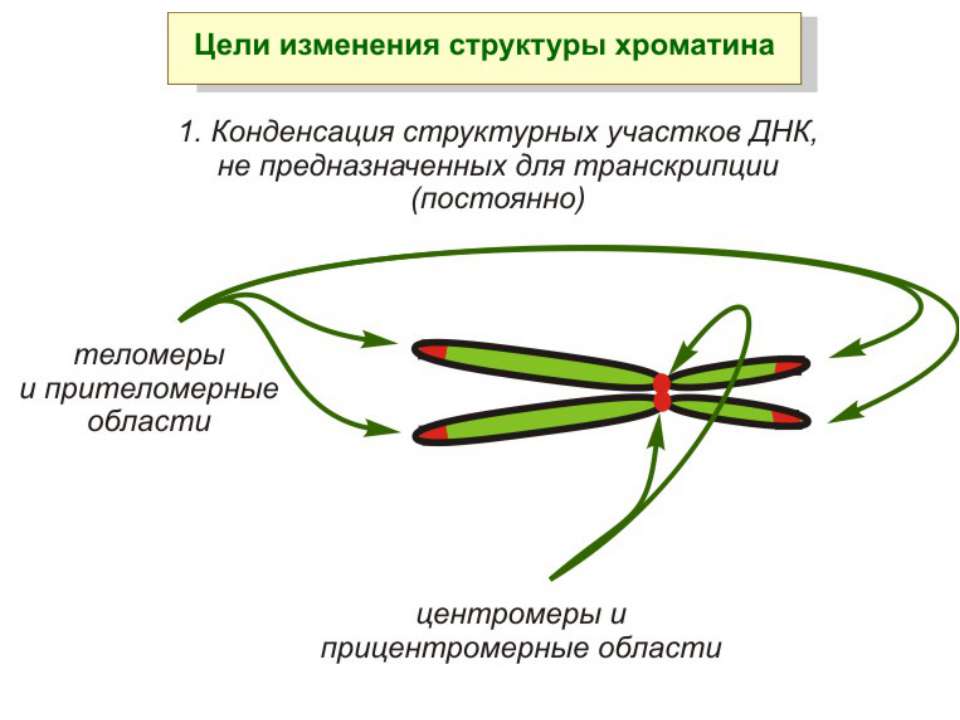
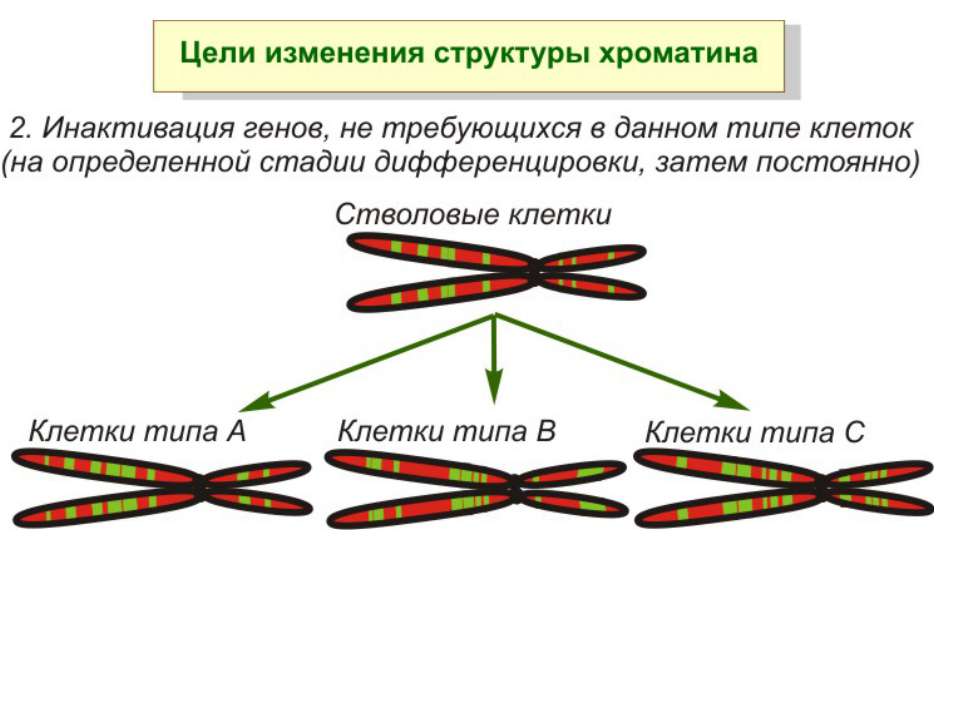
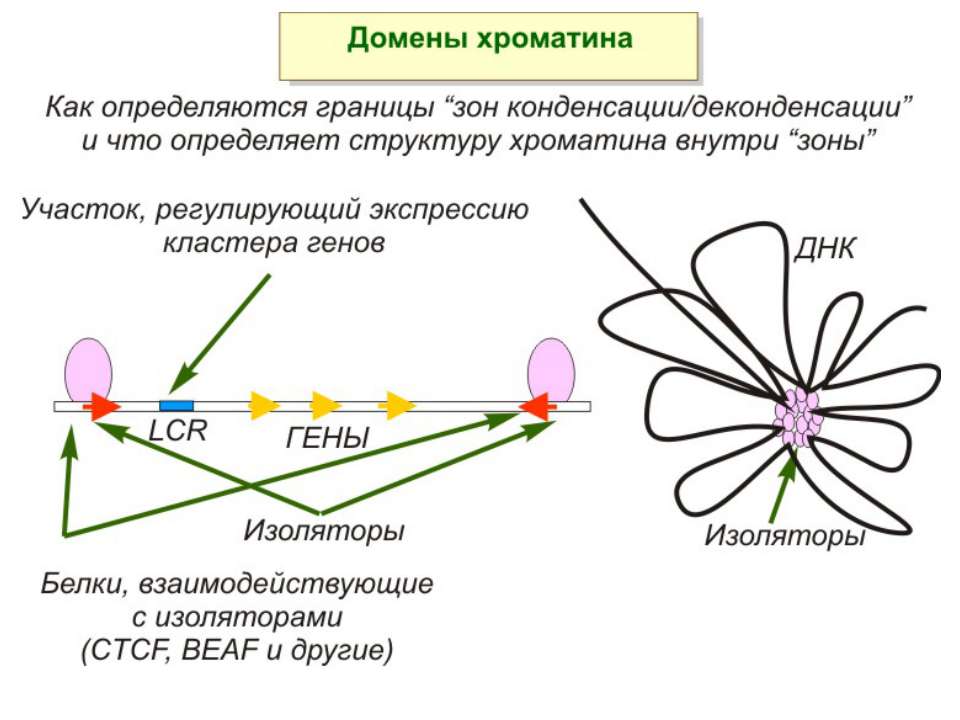
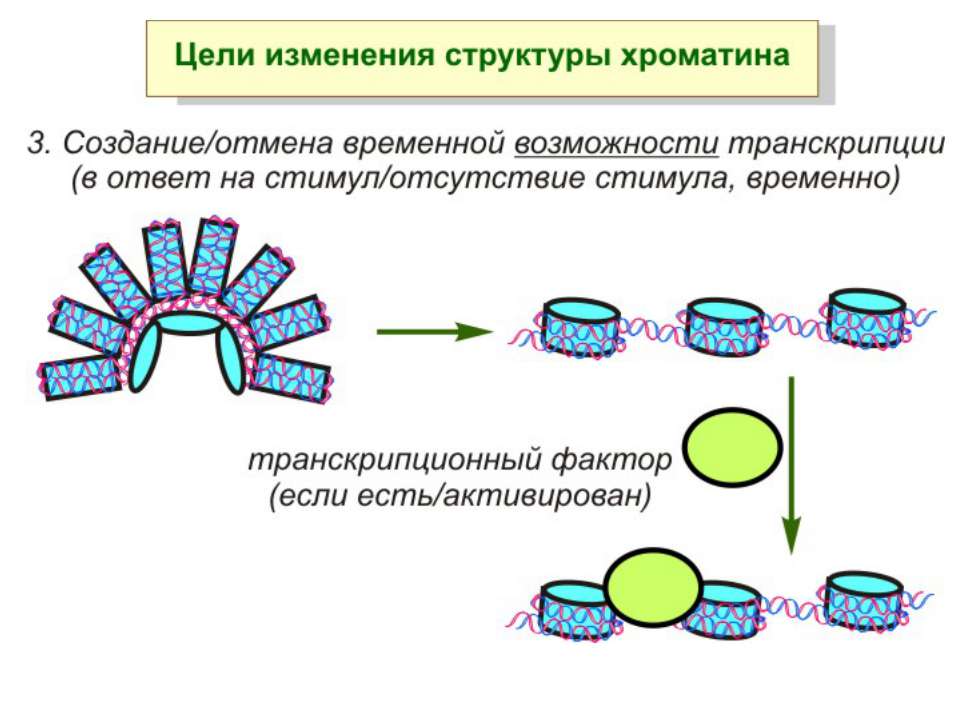
 ділянки ...")
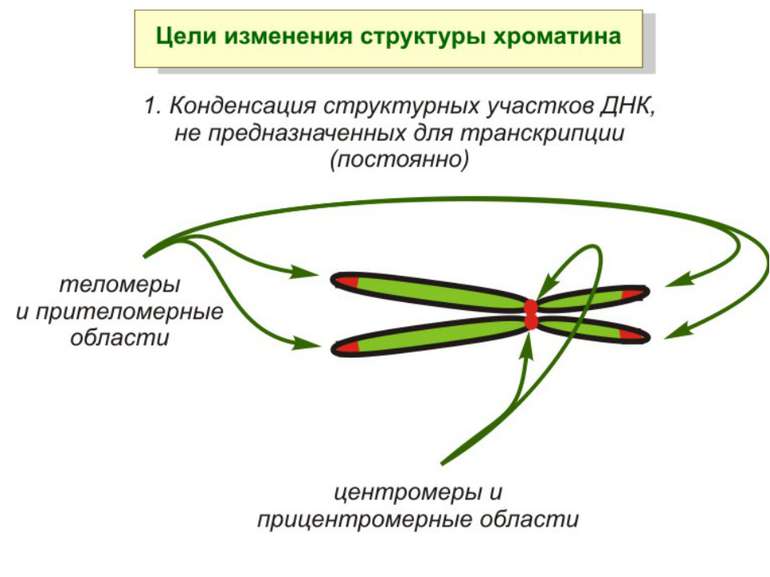
Хроматиновий рівень організації Великі (біля сотень тисяч пар основ) ділянки ДНК, що містять кілька генів - домени хроматину – утворюють випетлювання від осі хромосоми і можуть конденсуватись/деконденсуватись як єдине ціле Н.: глобінові гени
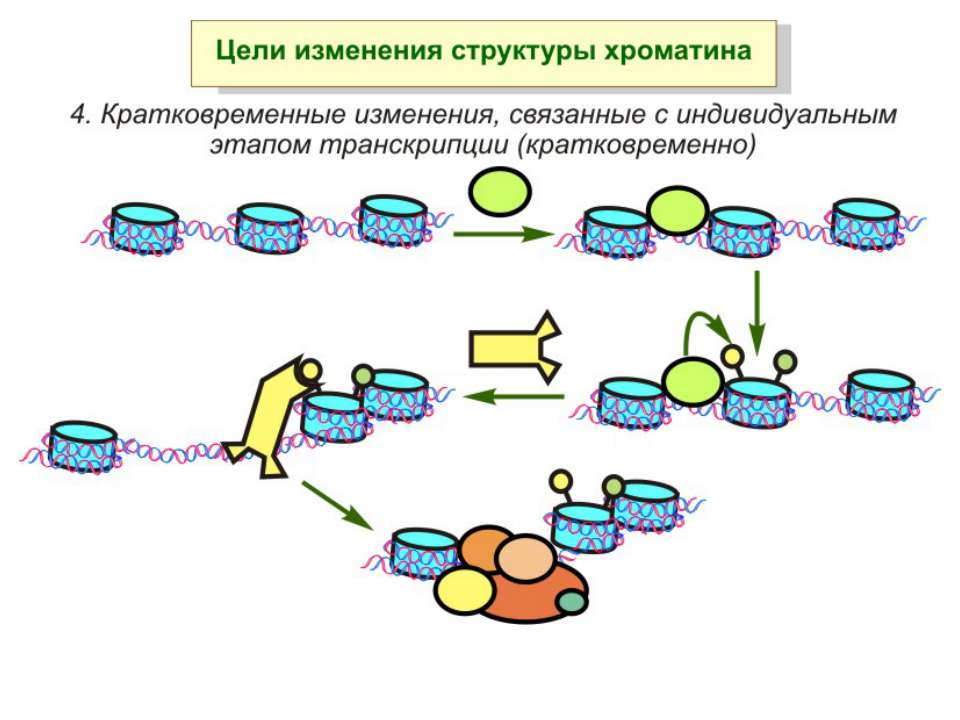

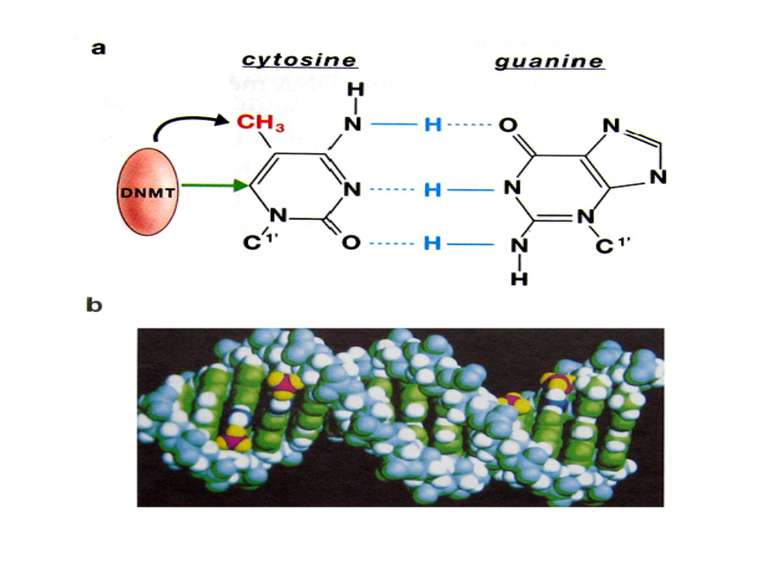
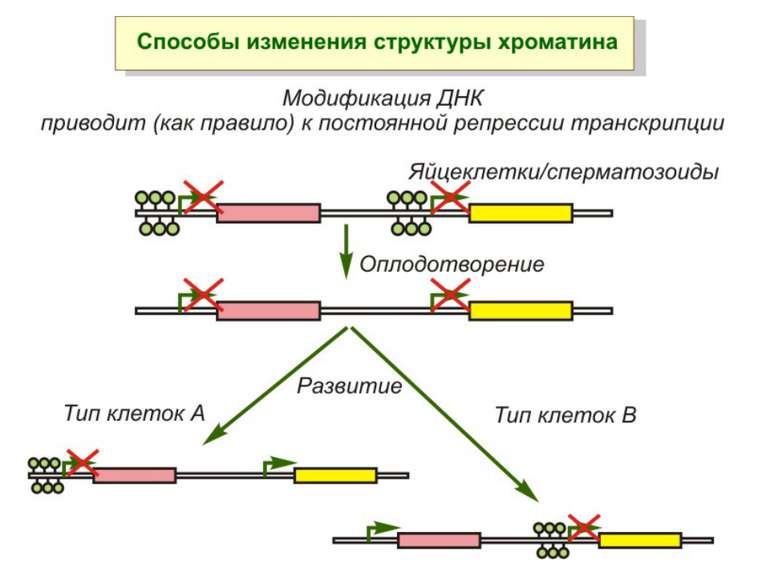
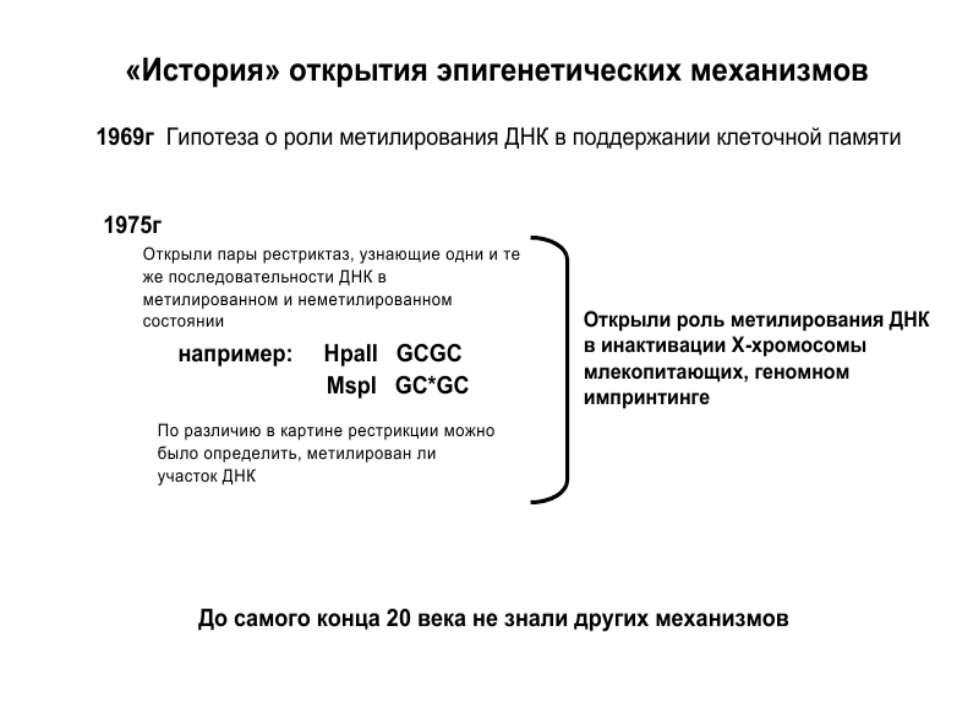
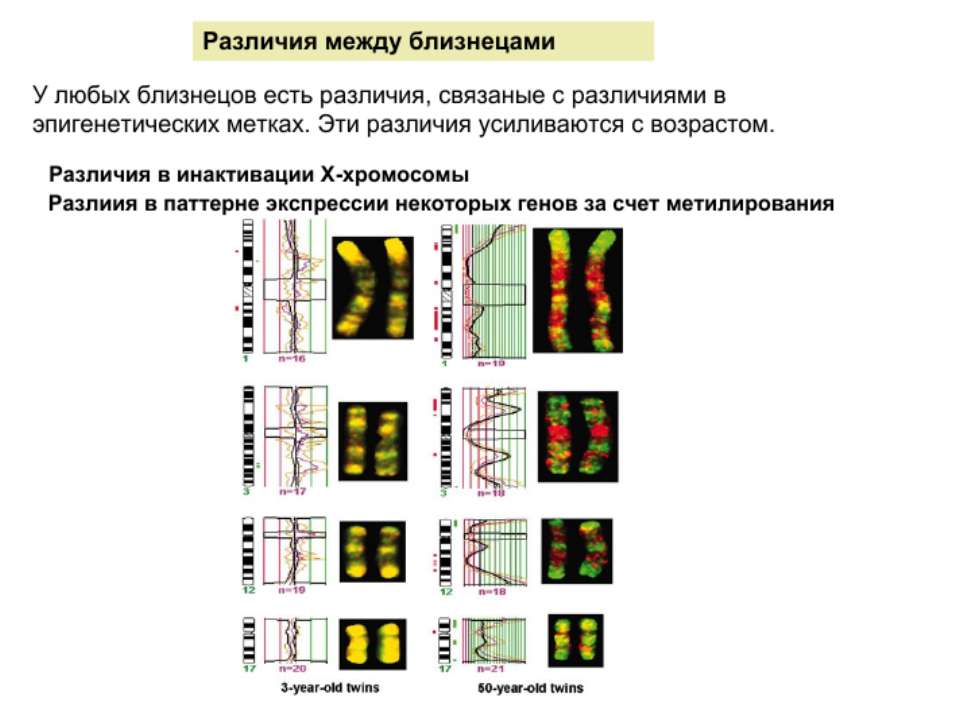
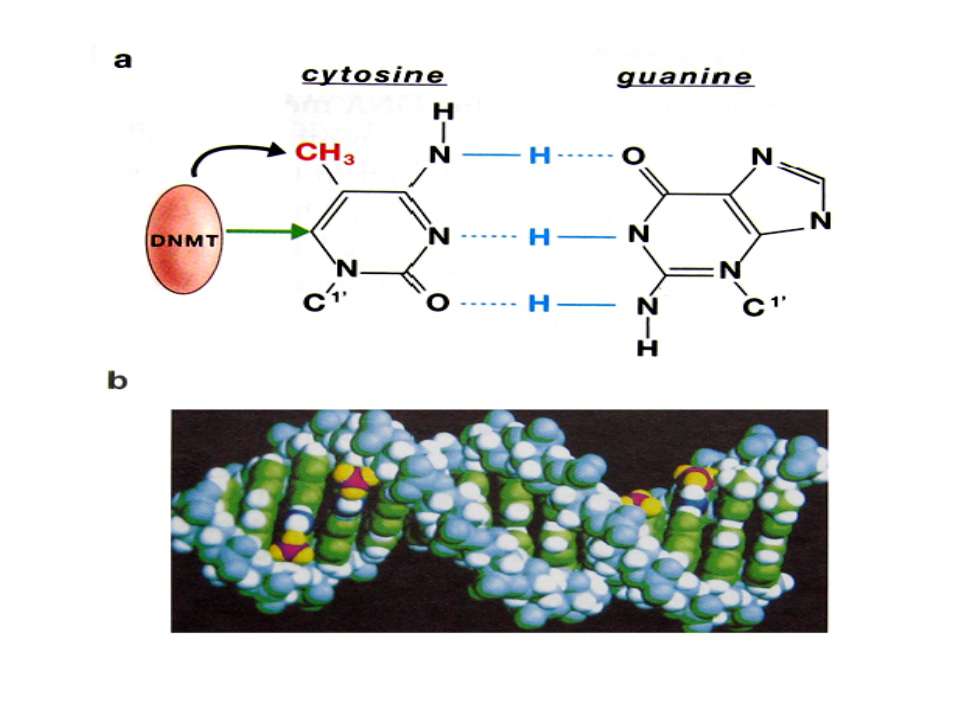
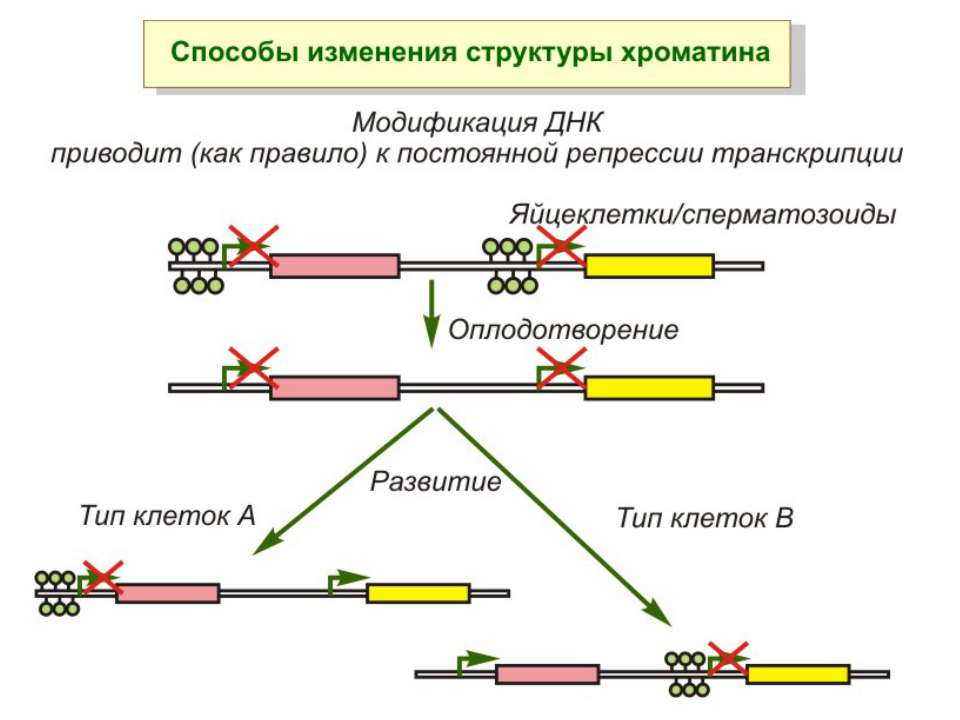
Роль метилювання ДНК в регуляції транскрипції Вибірковість метилювання Карти метилювання (батьківський імпрінтинг, хвилі онтогенезу) Метилювання і онкогенез Участь в інактивації Х-хромосоми (синдром Тернера)

mC Т !!!! С U Н.: мутації гена білка р53

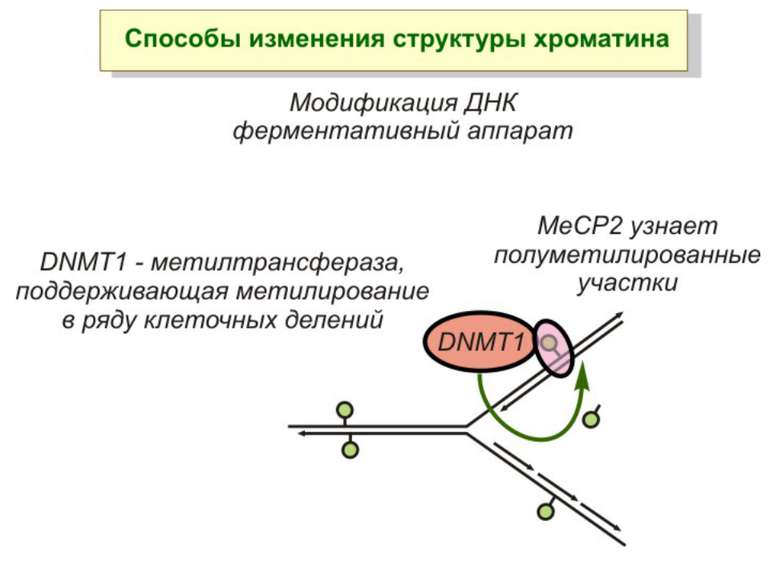
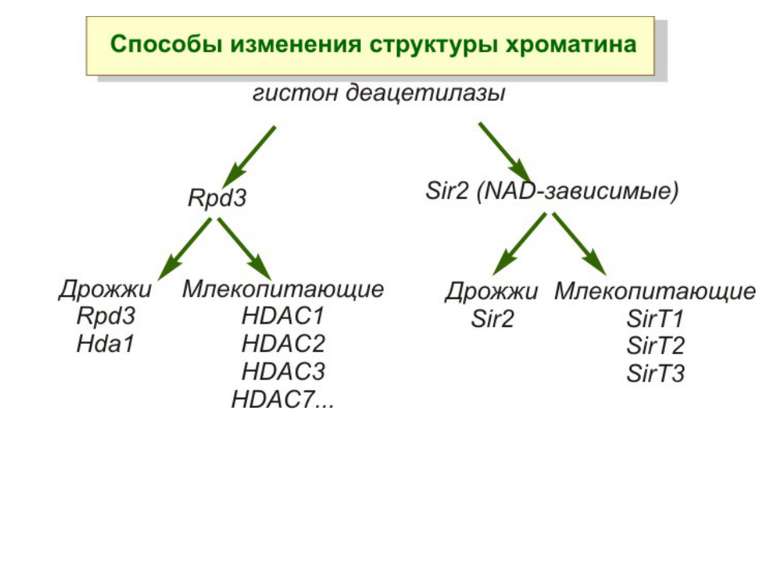
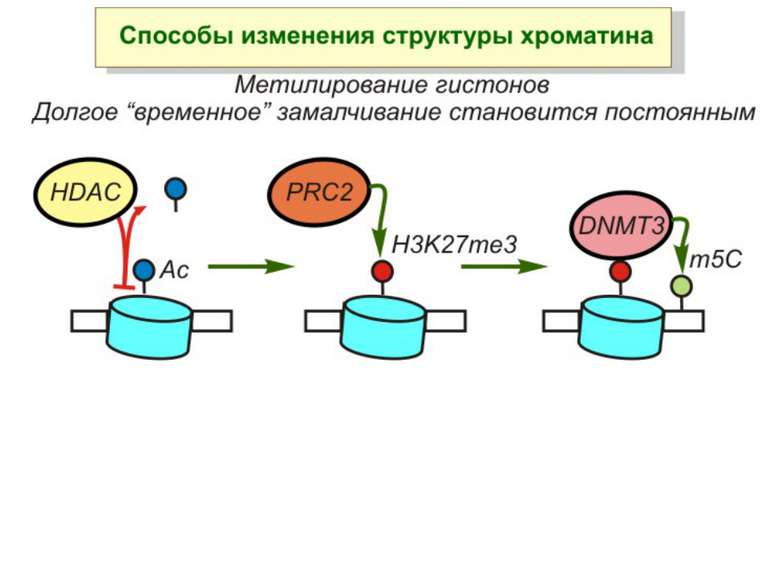
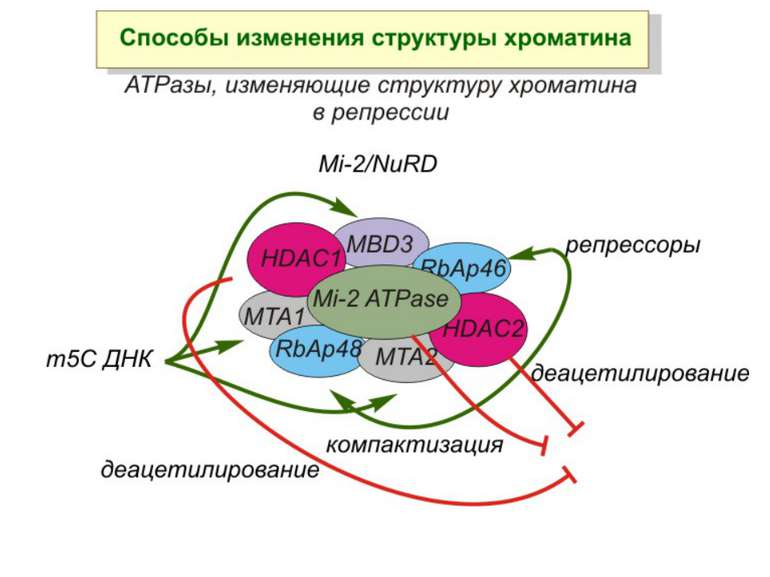
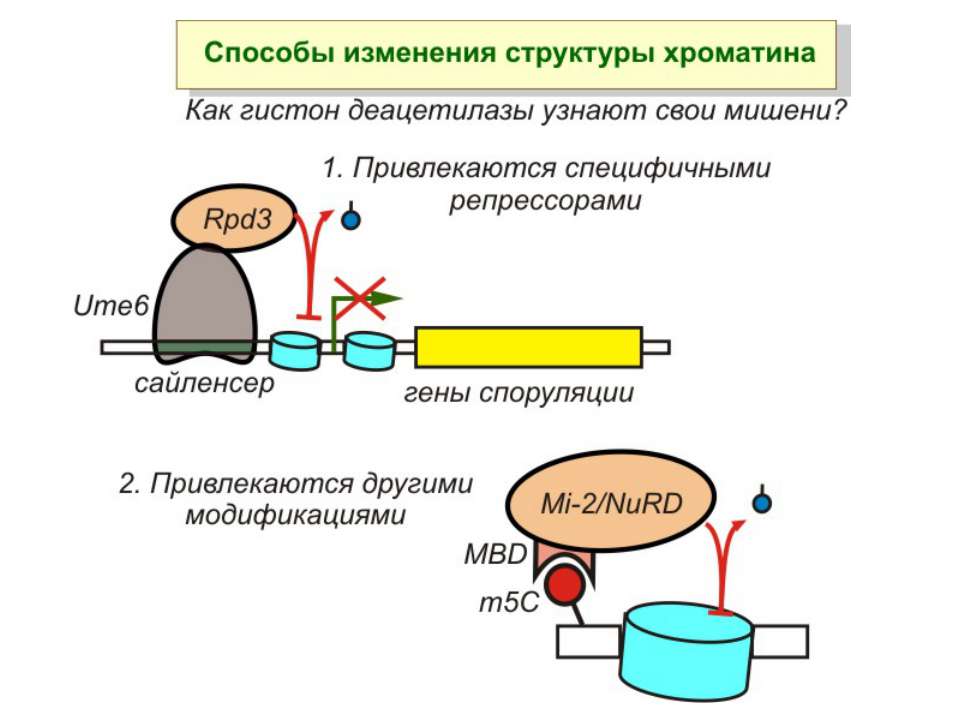
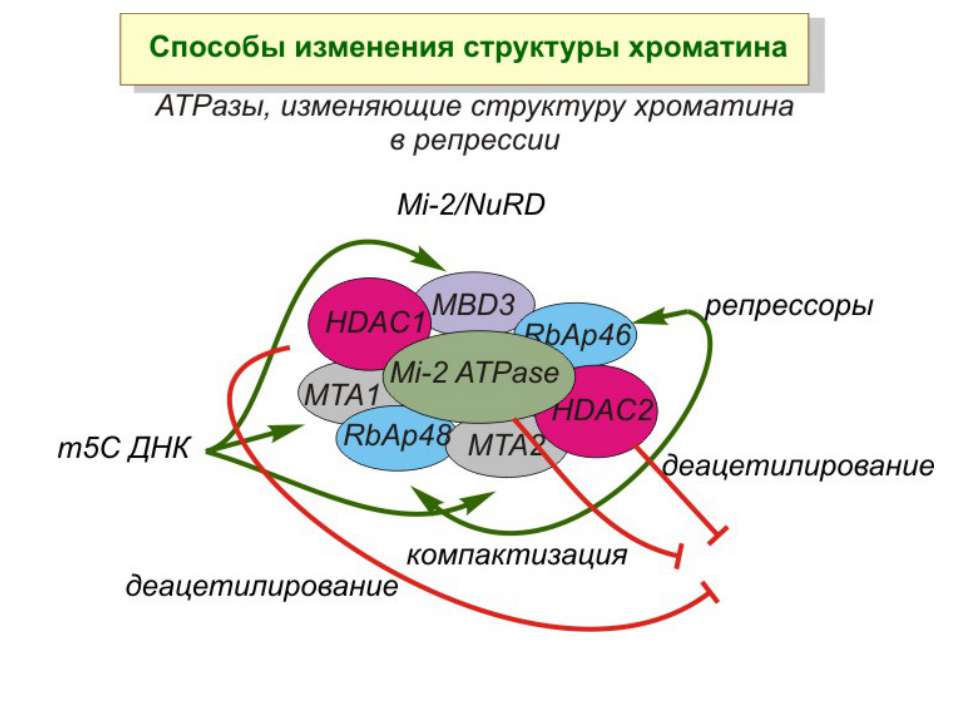
Зв’язок між метилюванням хроматину та конденсацією хроматину З метильованою ДНК зв’язуються білки, що містять MBD-домени. Ці білки, кооперативно з білками-репресорами можуть викликати посадку на ДНК складних білкових комплексів (Mi-2/NuRD), які забезпечують деацетилювання гістонів і активну, ATP-залежну конденсацію хроматину

NB!!! Метилювання інтронних ділянок – активація транскрипції

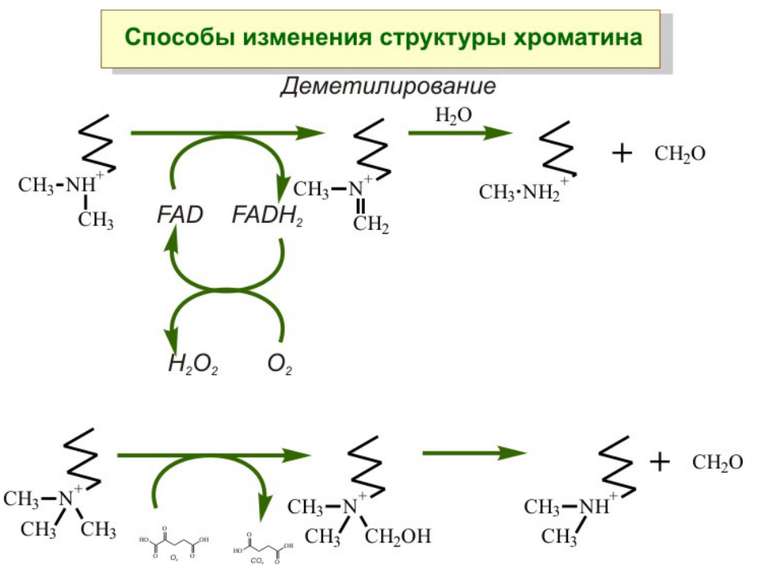
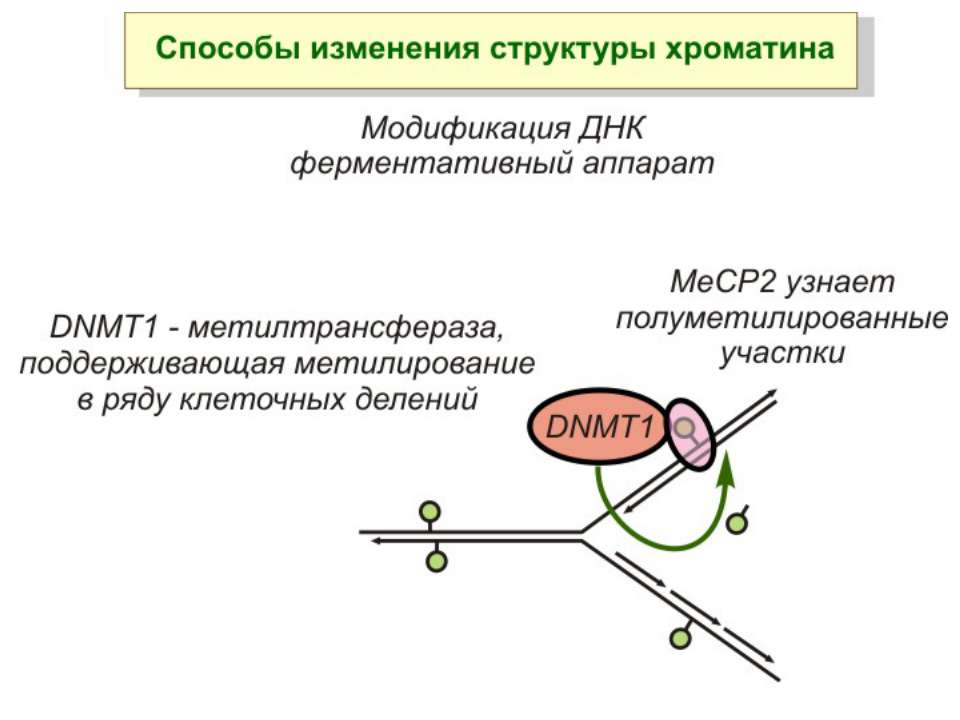
Передача карт метилювання Хвилі метилювання в онтогенезі (пасивне і активне деметилювання ((фактори транскрипції?))

Передача карт метилювання Батьківський імпритинг – сімейні спадкові захворювання аА* і аа Характерне для хребетних

Передача карт метилювання Еволюційне значення “Конфлікт інтересів” Ген фактору росту плаценти активний, якщо успадковується від батька, і гіперметильований (неактивний), якщо приходить від матері. Навпаки, ген, що інгібує фактор росту, активний лише в тому випадку, якщо успадковується від матері.

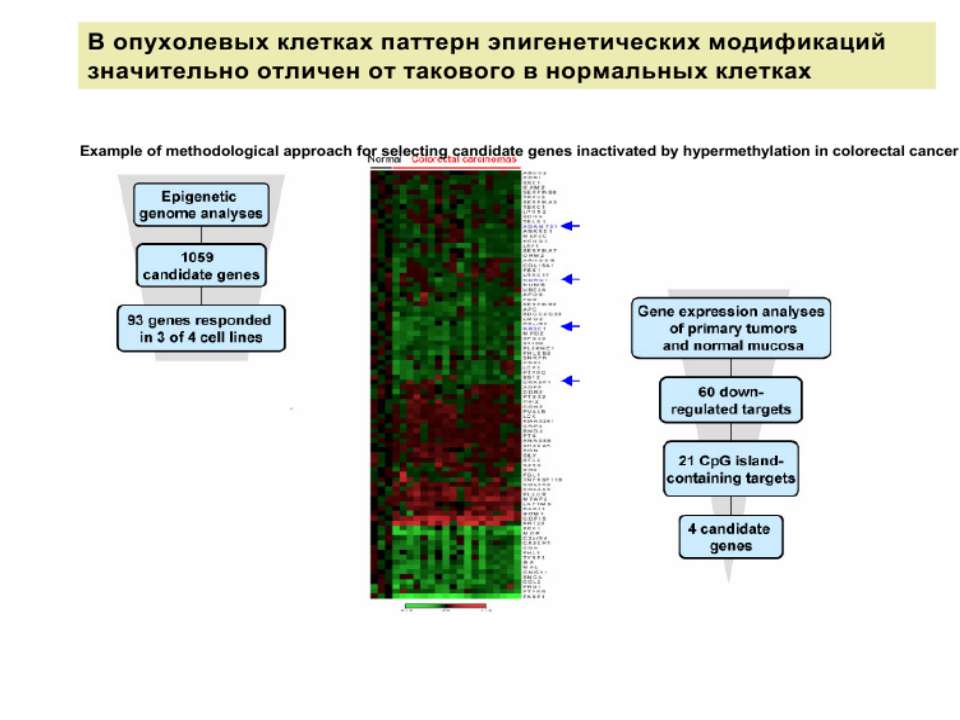
Метилювання і онкогенез Метилювання промоторів генів-супресорів Н.: р53 Rb (retinoblastoma)

Метилювання і інактивація Х-хромосоми Генний баланс – недопустимість дуплікацій аутосом Неповне виключення Випадковий вибір синдром Тернера – вплив походження Х-хромосоми

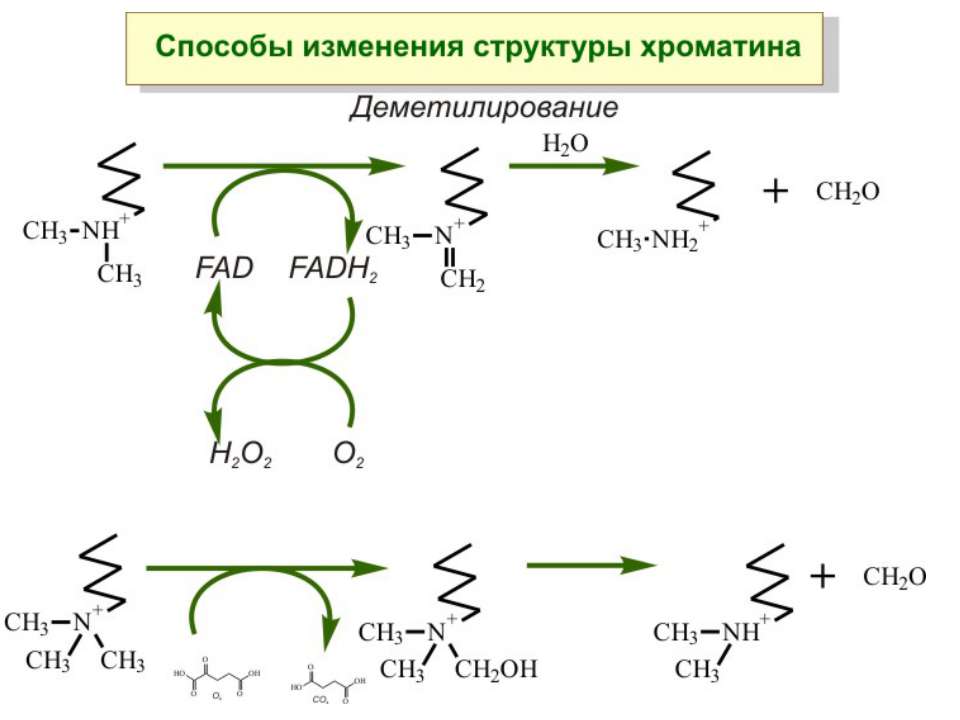
Метилювання аденінових залишків ДНК N6-метиладенін Залучене в регуляцію експресії генів і реплікацію, в тому числі і мітохондріальних ДНК. Може контролювати персистенцію чужерідних ДНК в клітині і, ймовірно, служить елементом системи рестрикції-модифікації у рослин.

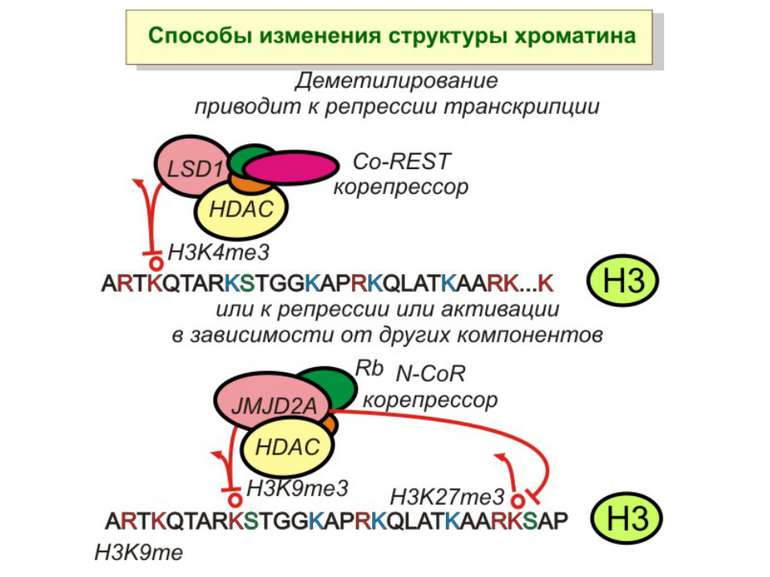
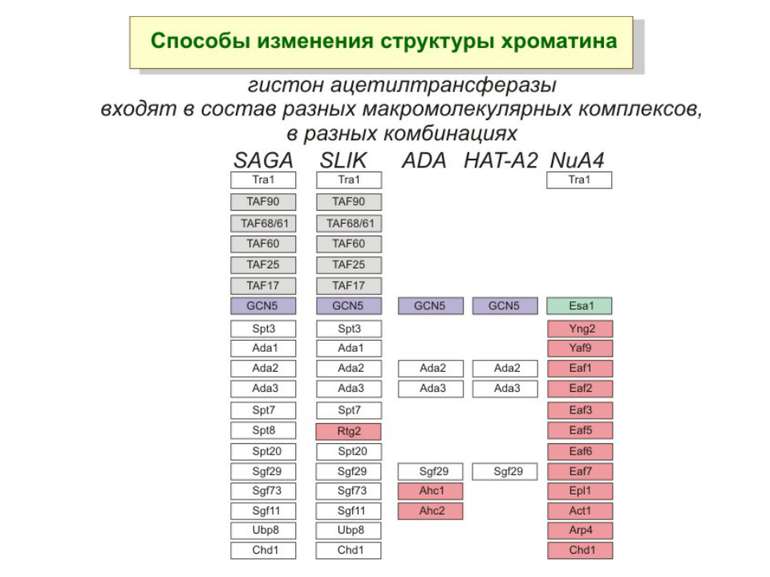
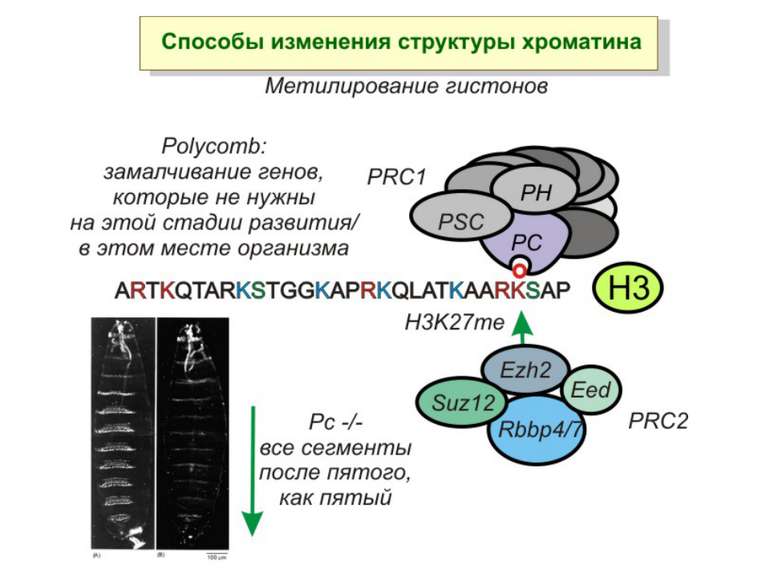
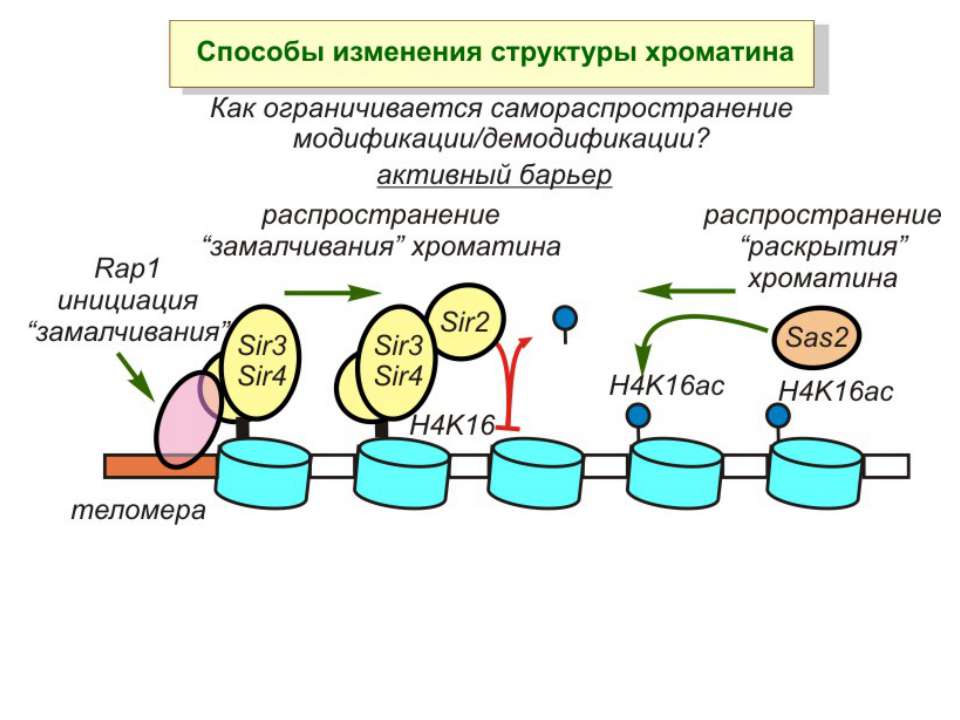
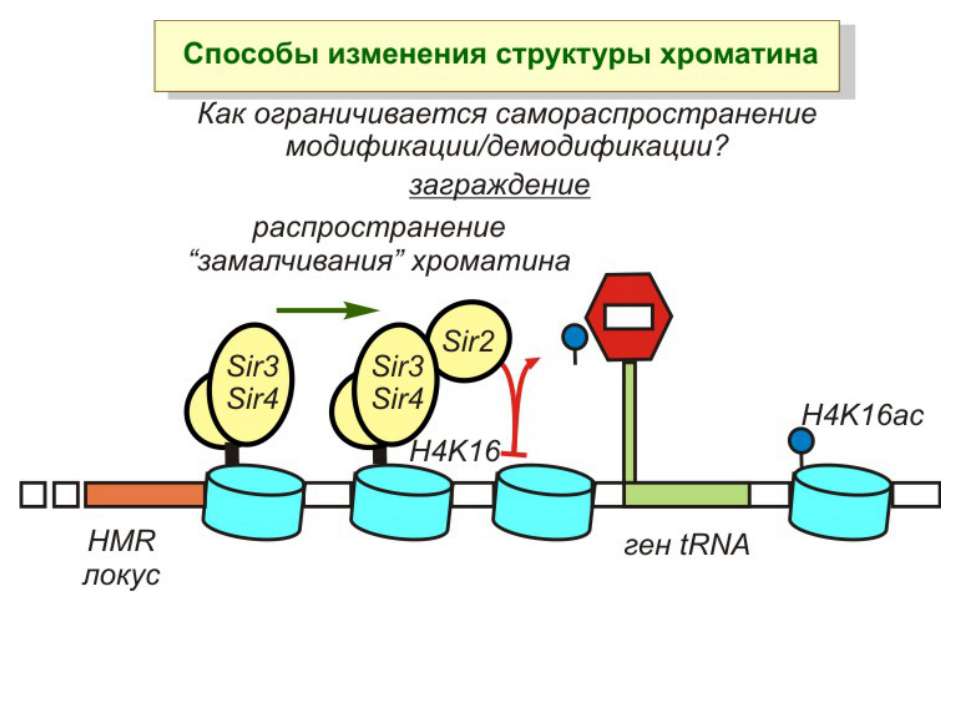
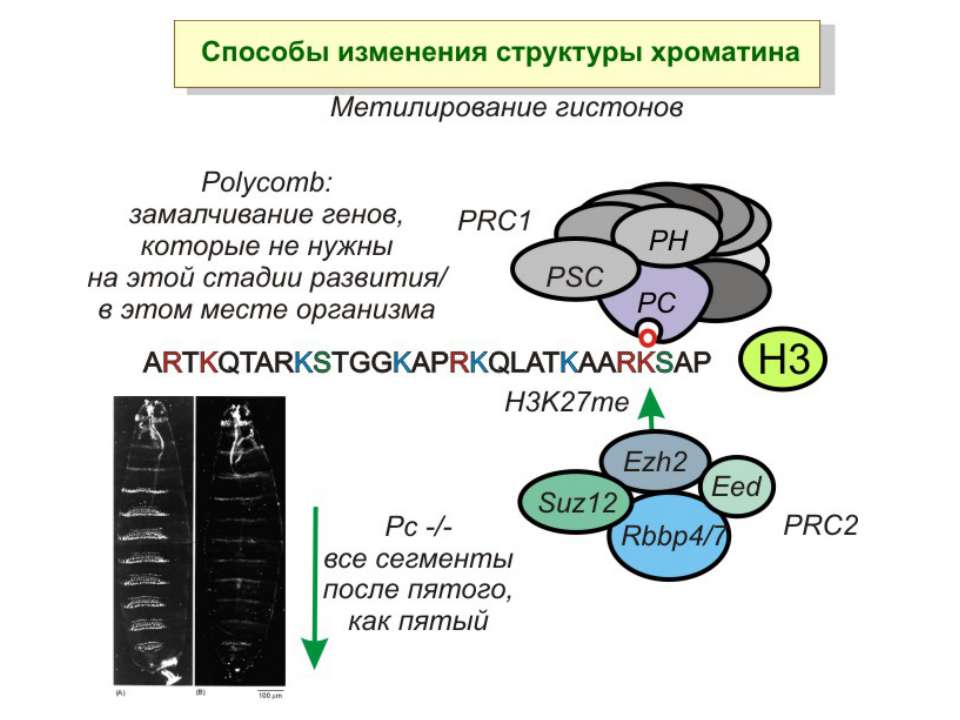
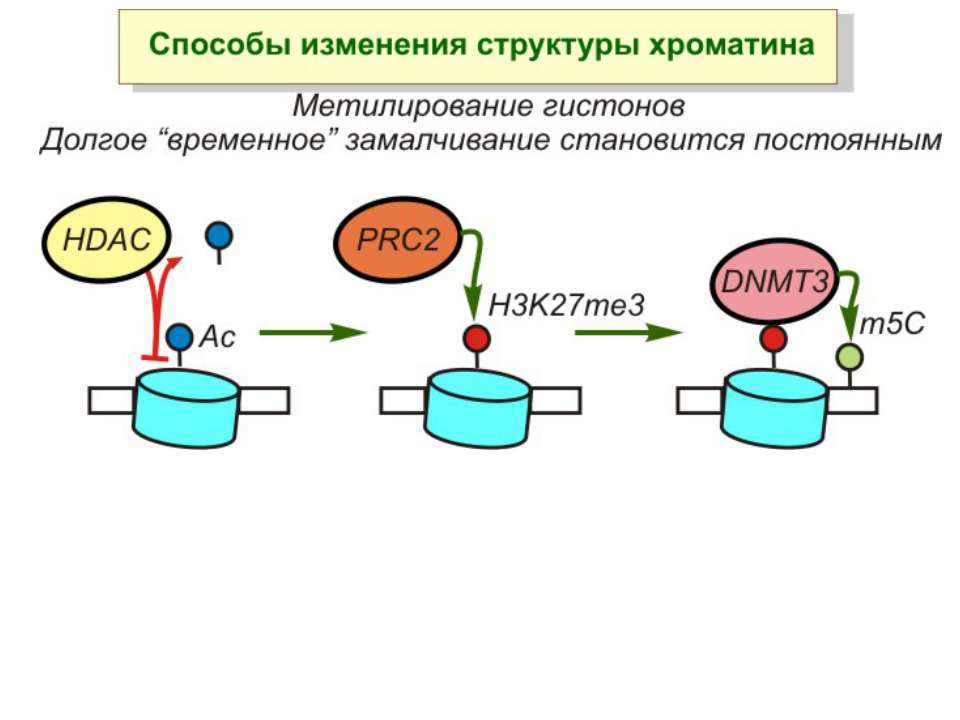
Способи зміни структури хроматину Модифікація гістонів високо консервативні лізини: гістона Н3 в положенні 9, 14, 18 і 23 гістона Н4 - 5, 8,12 і 16

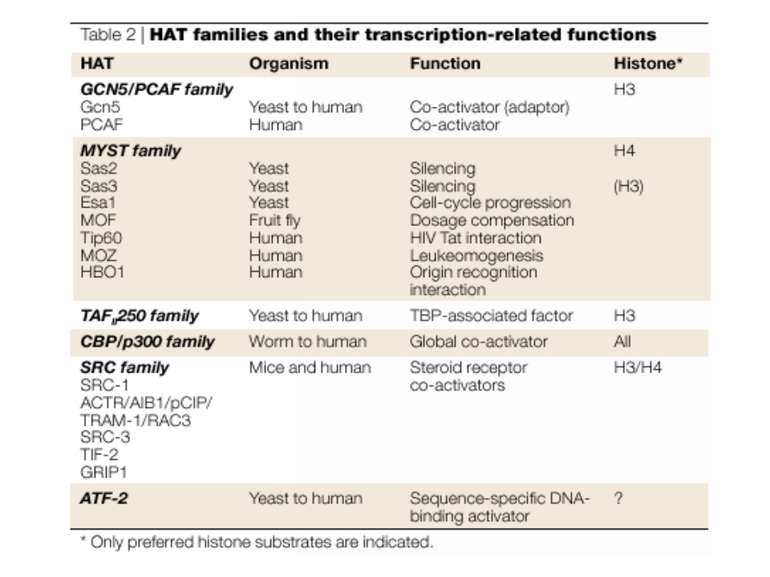
Protein modules that manipulate histone tails for chromatin regulation a | Mechanism of histone acetylation by HAT (histone acetyltransferase) proteins. The acetyl moiety is transferred from acetyl-coenzyme-A to the target lysine on the histone substrate. b | Structure of the Tetrahymena Gcn5–CoA–histone-H3 peptide complex. The CoA cofactor is shown in red and the histone H3 peptide is shown in green. The central core domain responsible for CoA binding and catalysis is blue; the amino- and carboxy-terminal subdomains associated with histone H3 binding are purple; and regions showing no sequence or structural homology with other HAT proteins are aqua. c | Structure of the MYST HAT family member yeast Esa1, bound to CoA58. Colour coding is the same as in (b).
 - ферменти, що епігенетично модифікують г...")
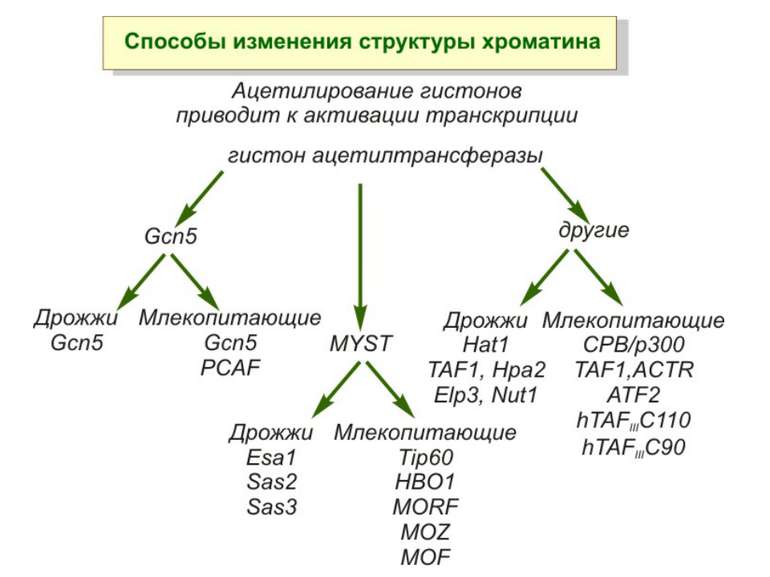
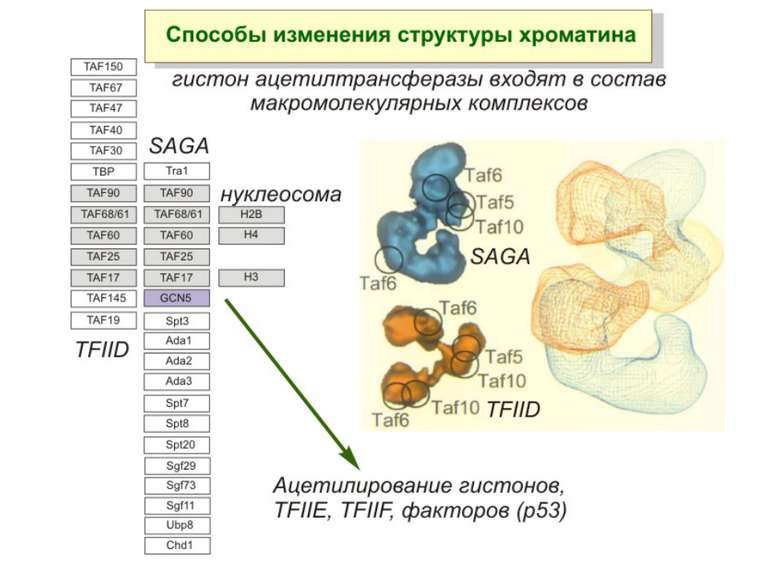
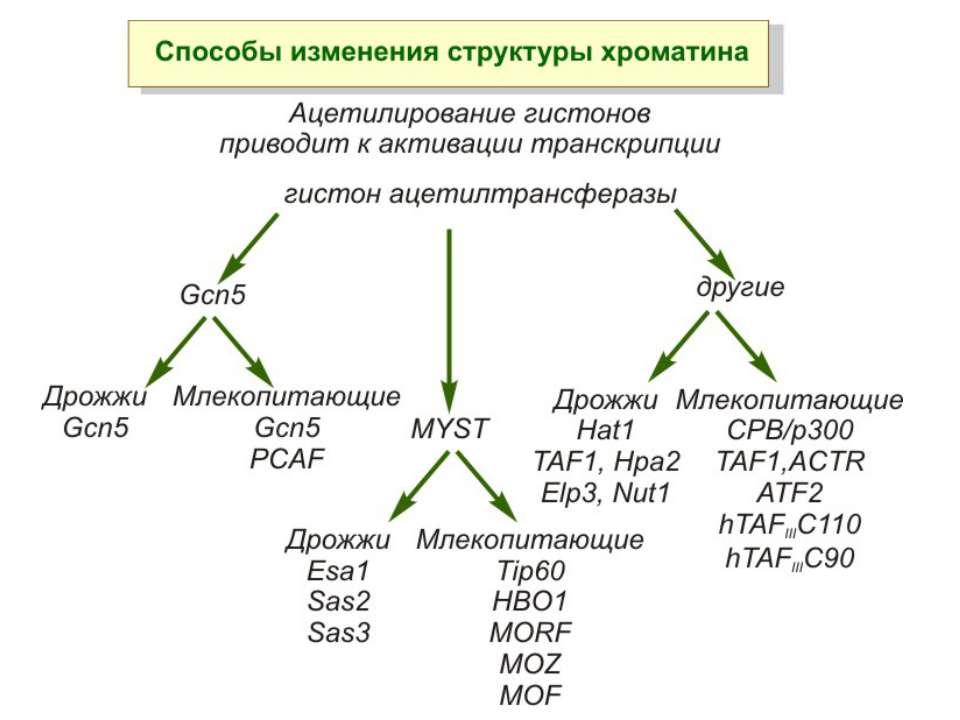
HAT Гістон ацетилтрансферази (Hats) - ферменти, що епігенетично модифікують генну регуляцію. Те, як вони модифікують свою мішень, залежить від їхньої форми. Ця кільцева структура утворено двома ацетилтрансферазами Rtt109 (показані фіолетовим і золотавим кольором і двома шаперонамиVps75 (синій і зелений) - білками, що супроводжують ферменти до їхньої мішені й, частково, визначають розмір отвору, що утворюється при формуванні кільця . Яка частина мішені модифікується, визначає форма кільцевого комплексу, що утворюється , коли HAT зв'язується з верхньою частиною гістонового білка. Інші шаперони можуть допомогти Rtt109 модифікувати інші частини гістона.
 включає білки, що беруть участ...")
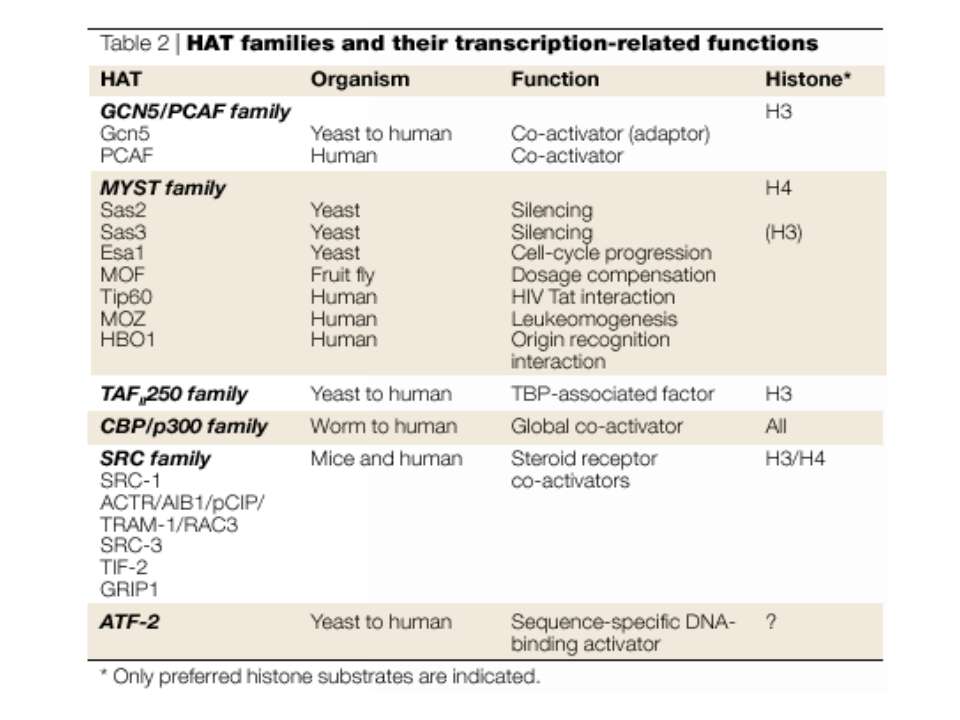
GNAT родина (Gcn5-related N-acetytransferases) включає білки, що беруть участь або зв’язані з ініціацією транскипції (Gcn5 і PCAF), елонгацією (Elp3), депозицією гістонів і теломерним мовчанням (Hat1)

Родина p300/CBP HAT Наявні спільні гомологічні послідовності з GNATs. HAT активність p300 і CBP необхідна для їх роли в трансактивації, вони асоціюють з ін. ацетилтрансферазами. P300/CBP - HAC, взаємодіють з енхансерами і асоціюють з РНК - полімеразою II
, Ybf2/Sas3, Sas2 і Tip60. Н.:...")
Родина MYST Включає членів MOZ (онкоген людини), Ybf2/Sas3, Sas2 і Tip60. Н.: гостра мієлома Участь в транскрипційній елонгації і реплікації та в мовчанні дріжджевих локусів HM типів спарювання. На відміну від SAGA, комплекс NuA3 не рекрутується дріжджевими промоторами за допомогою кислих активаторів.
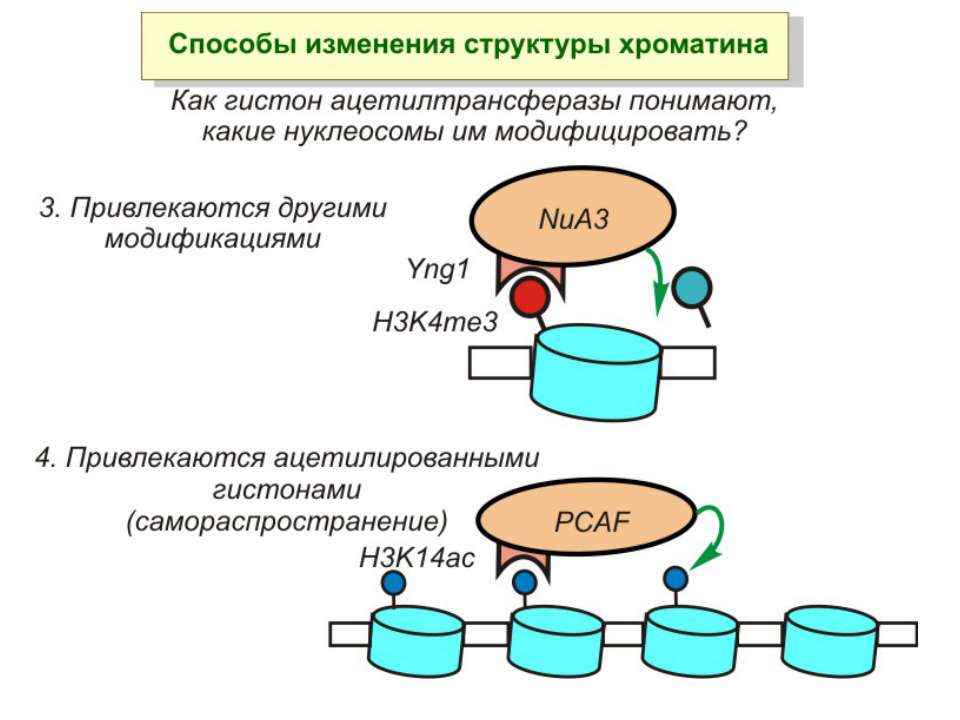



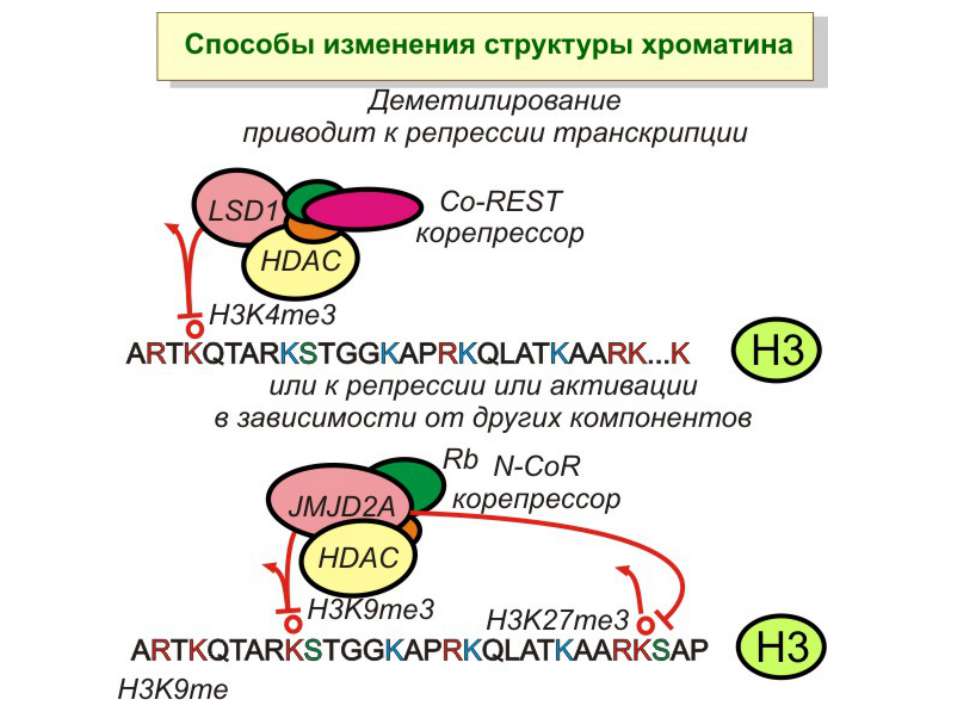
в гістонах переважно метилюються в H3 лізини 4, 9 і 27, а в H4 лізин 20 Модифіковані лізини можуть бути моно-, ди- чи три- метильованими

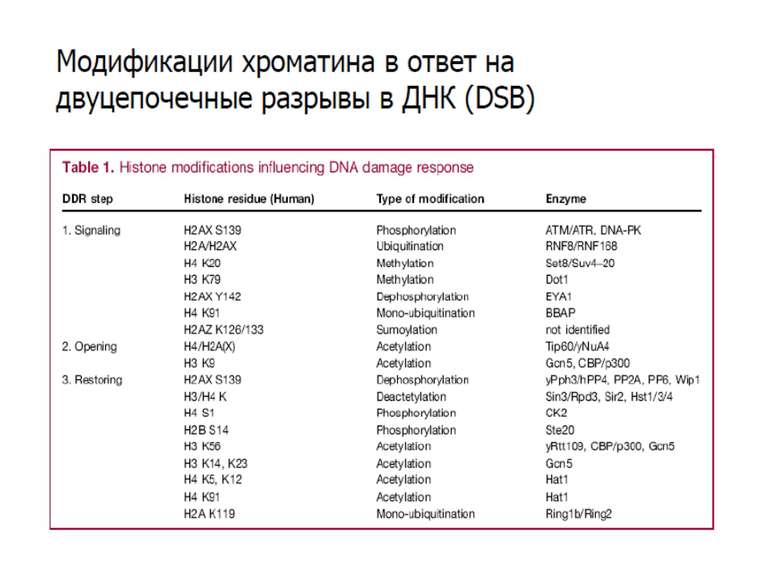
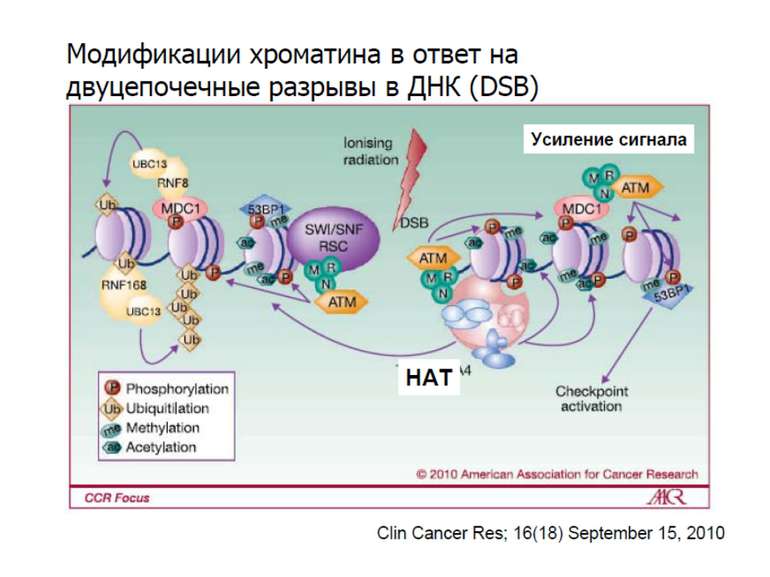
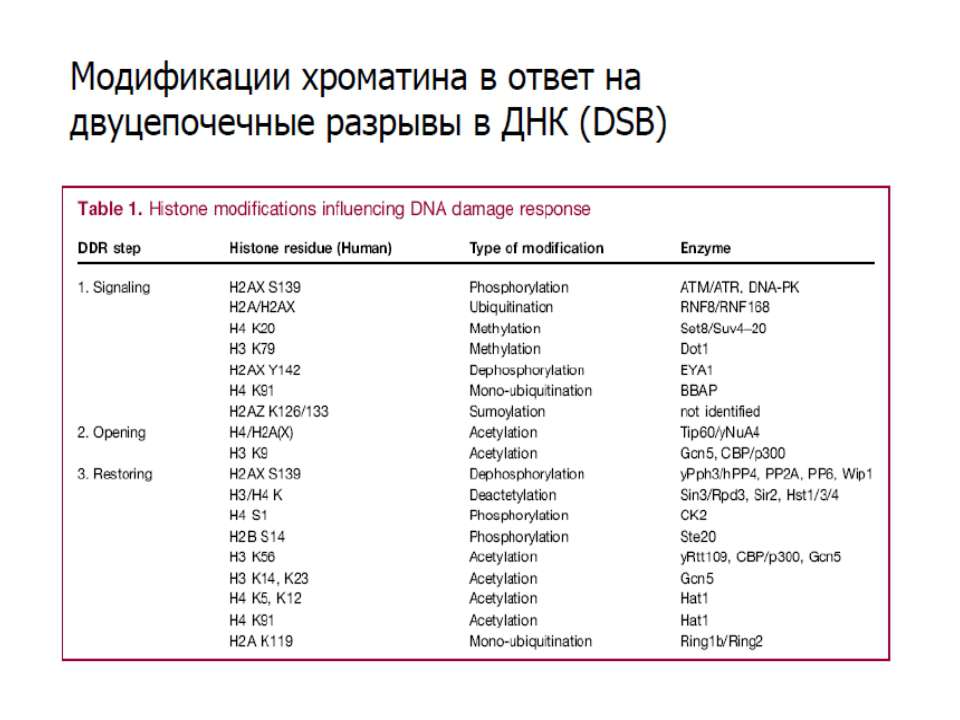
Фосфорилювання гістонів Тісно спряжене з ацетилюванням Н.: Ліз-9 Сер-10 Може відбувати у відповідь на пошкодження ДНК (ASQE-мотив Н2А)

NB !!! Метилювання і ацетилирювання Lys9 взаємовиключаючі. Метилювання Lys9 інгібує фосфорилювання Ser10, а не ацетилювання Lys14. Метилювання Lys9 корелює з мовчанням генів, тоді як фосфорилювання Ser10 і ацетилювання Lys9 корелює з транскрипційною активністю

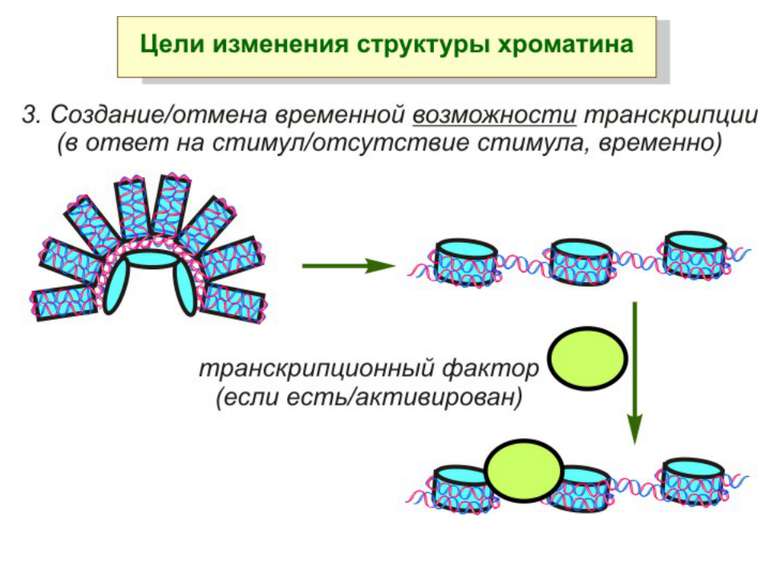
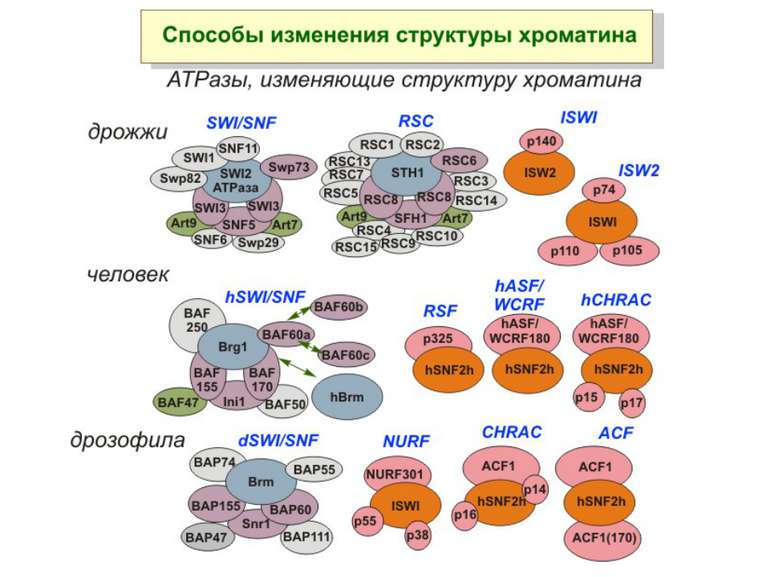
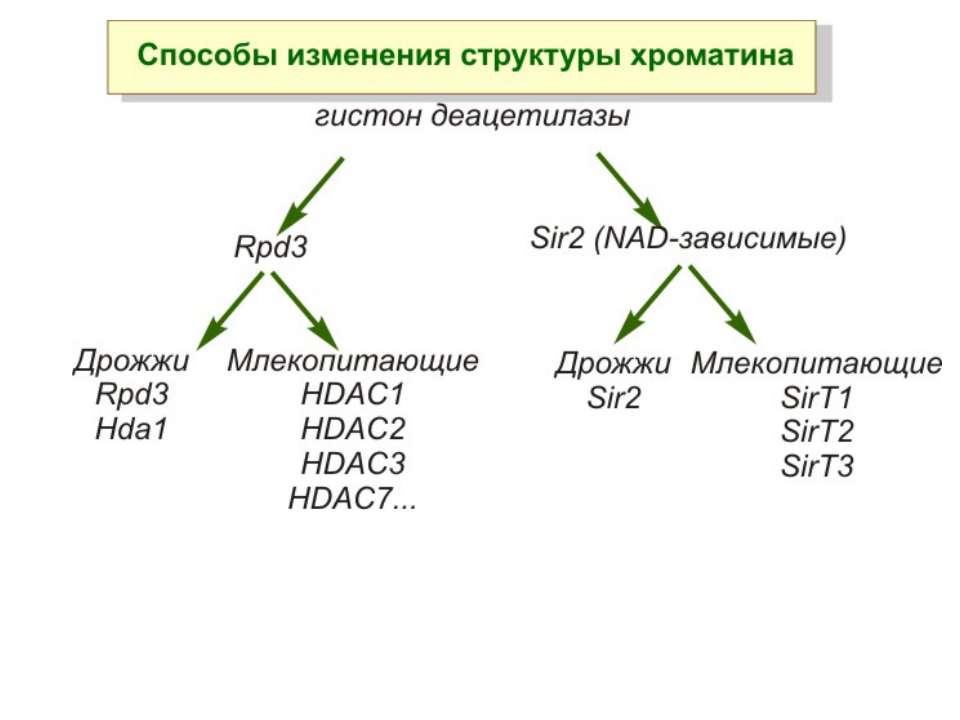
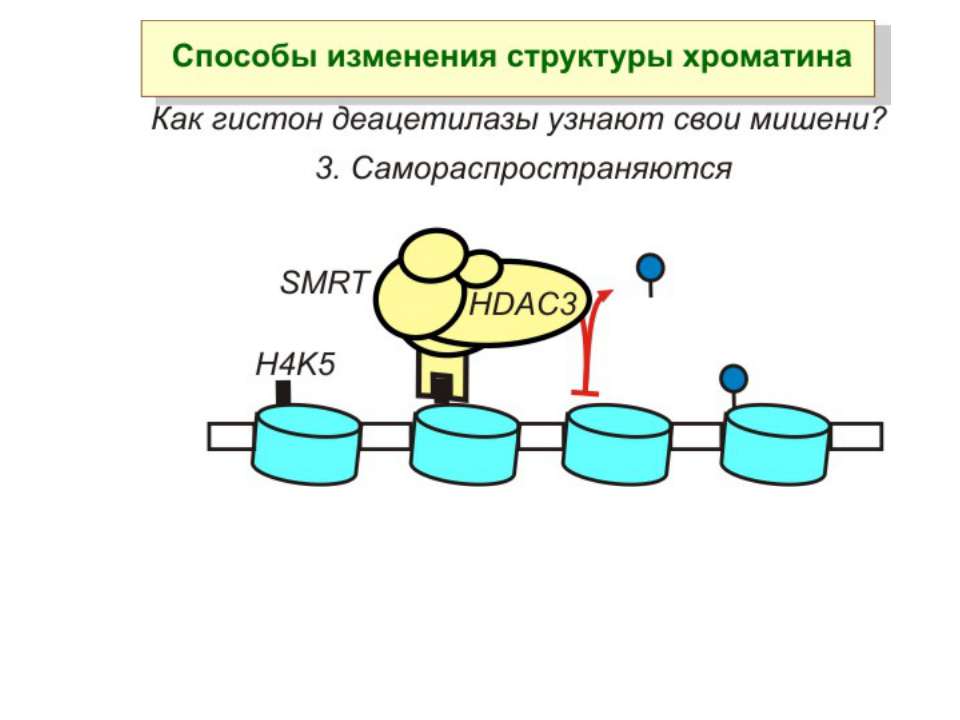
Зміщення нуклеосом
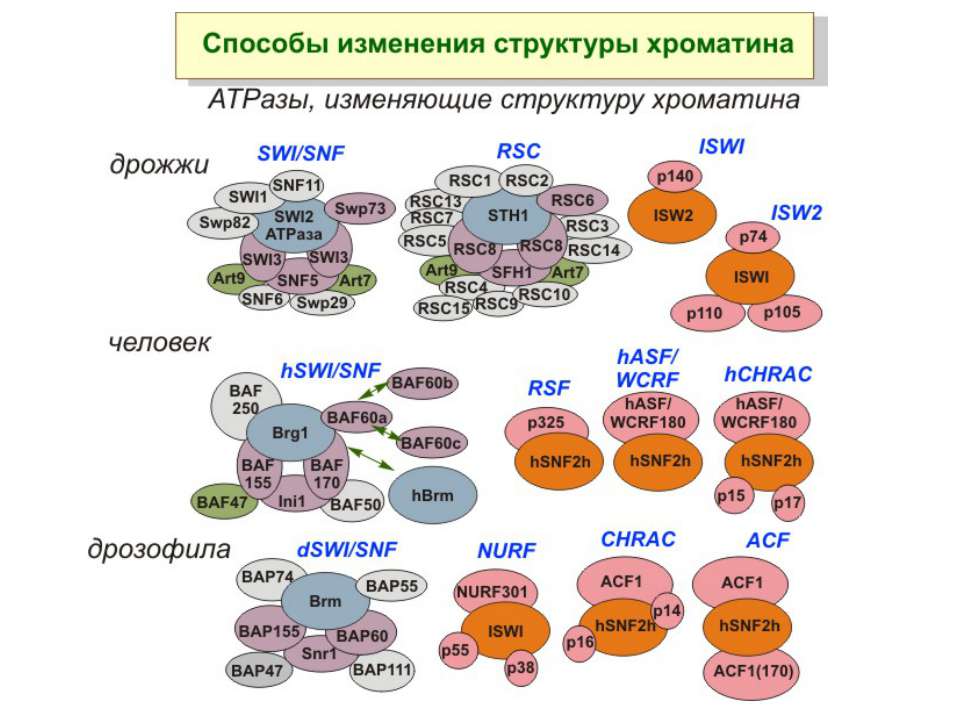


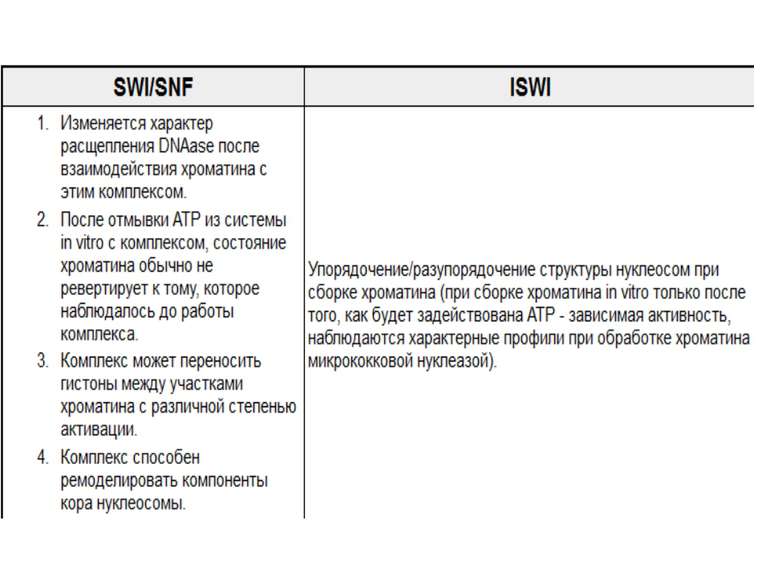
Комплекси, що містять гомологи SWI2 і ISWI відрізняються !!! SWI2 може використовувати як субстрат ненуклеосомну ДНК, і може виконувати складні перебудови ДНК-гістонового комплексу. Субстратом ISWI жуть бути лише нуклеосоми, а основною функцією – зсув нуклеосом по ДНК. Причому для ISWI необхідним є N-кінець гістону H4.
 визначається їх функцією")
Місцезнаходження гістонів (та їх варіантів) визначається їх функцією

Де знаходяться гістони ? Послідовності ДНК можуть в 1000 раз відрізнятися по потенціалу зв'язувати нуклеосому Це пов'язане з анізотропною гнучкістю ДНК. Пари нуклеотидів AG/CT, CG/GC, GA/TC, GC/CG, AA/TT згинають ДНК Якщо підряд ідуть послідовності, що згинають ДНК в одну сторону (наприклад, TATA!), зв'язування нуклеосоми буде нестійким. Протилежно спрямований вигин ДНК із періодом, що відповідають 1 витку спиралі ДНК – максимально міцне зв'язування з нуклеосомою . Знаючи тільки первинну структуру ДНК можна передбачити точне положення більш 50% нуклеосом! («A genomic code for nucleosome positioning»)

Ідеальна послідовність

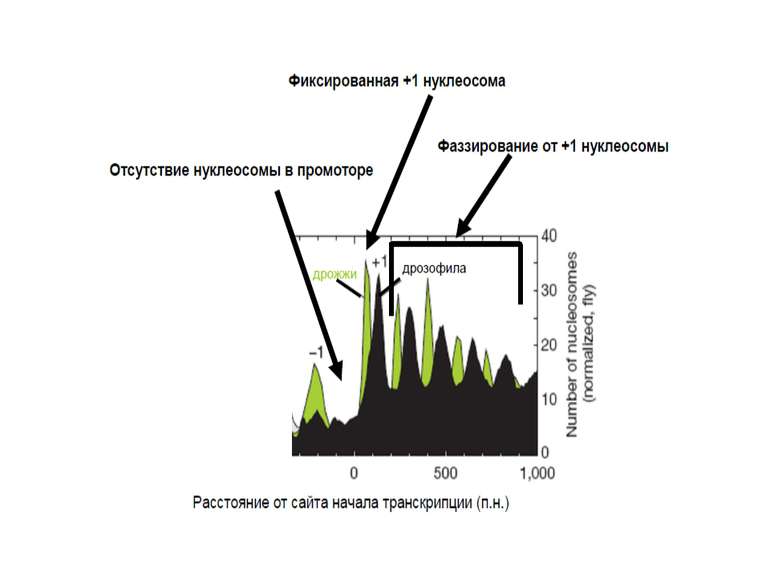
У геномі присутні Ділянки, вільні від нуклеосом сайти зв'язування транскрипційних факторів, регуляторних білків Ділянки, де положення нуклеосоми строго фіксоване +1 нуклеосома в генах (дріжджі – від +1 нуклеотиду, хребетні – від +60) Ділянки, у яких нуклеосомна укладка піддається регуляції білками АТФ-залежного ремоделінгу хроматину Ділянки, у яких спостерігається фазування нуклеосом, починаючи від фіксованої нуклеосоми
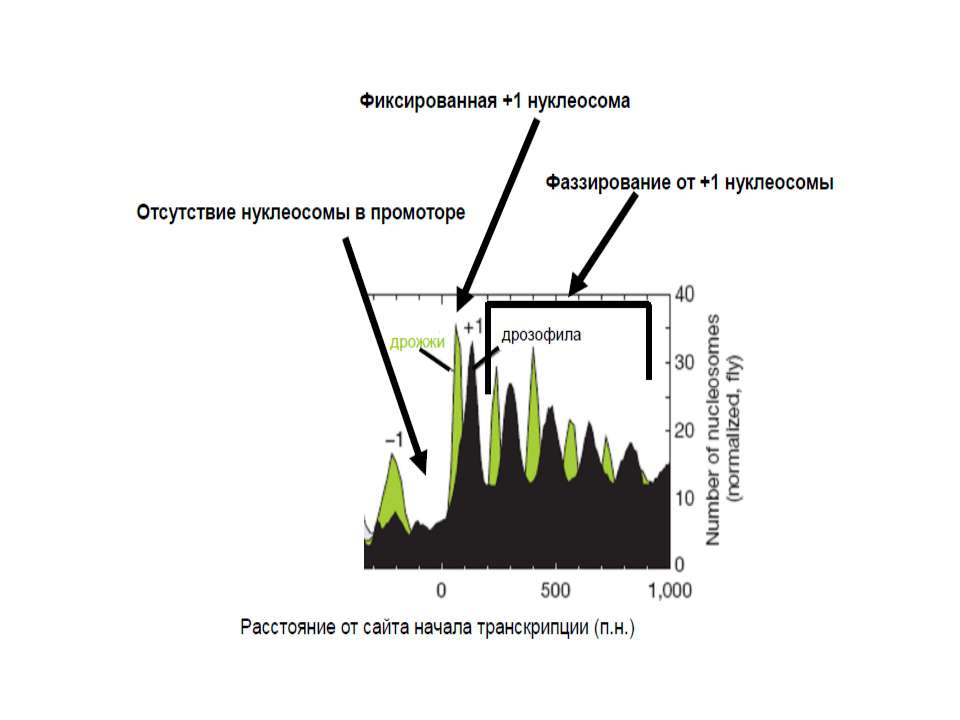

У промоторах майже усіх генів дріжджів поблизу від старту транскрипції наявна ділянка, вільна від нуклеосом

Стабільність зв'язування між ДНК і нуклеосомою може змінюватися при заміні основних варіантів гістонів на специфічні й у результаті ковалентних модифікацій гістонових хвостів Варіанти гістонів кодуються окремими генами. Вони гомологічні генам основного набору гістонів, але менш консервативні

Варіанти гістона Н3

Варіанти гістона Н2А

Різні гістони – різні модифікації

Варіанти гістонів розподілені на хромосомі у залежності від їх функції

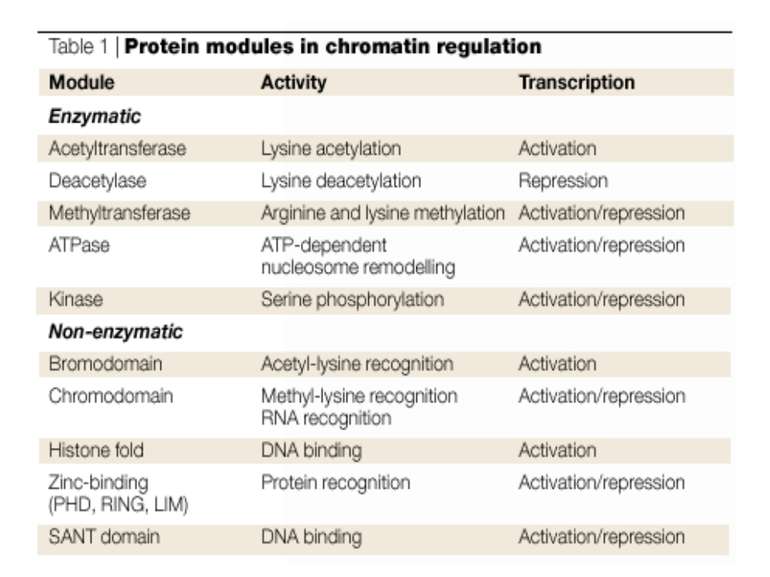
Посттрансляційні модифікації гістонів – “молекулярні мітки” Неепігенетичні Епігенетичні Успадковуються “гістоновий код”

“Гістоновий код” різні паттерни модифікації гістонів (результат ацетилювання, фосфорилювання, метилювання і ін. пост-трансляційних модифікацій) діють послідовно або в комбінації для формування «гістонового коду», який зчитується за допомогою інших білків, щоб викликати певні низхідні транскрипційні події.

Як працює “гістоновий код” 1. Амінокислотний залишок у гістоні 2. Модифікуючий фермент 3. Білок, який “сприймає” модифікацію

Домени білків, що розпізнають метилювання в лізині, ацетилювання в лізині, фосфорилювання в серині Ацетильований лізин - bromo-domain Метильований лізин - chromo-domain Tudor-domain

Protein modules that manipulate histone tails for chromatin regulation Representative members of different chromatin-regulatory proteins are shown with their associated modules. Bromo, bromodomain; Chromo, chromodomain; Chromo-S, chromoshadow domain; Cys, cysteine domain; HAT, histone acetyltransferase domain; Kinase, kinase domain; SET, Su(var)3-9, Enhancer of zeste, Trithorax; TAZ, PHD and ZZ domains are conserved cysteine–histidine-rich regions; Zn, zinc-binding domain

Structure of the bromodomain module a | Structure of the yeast Gcn5 bromodomain in complex with a histone H4 peptide substrate. The lysine and acetyl moieties of acetyl–lysine are indicated in red and green, respectively. b | Surface representation of the TAFII250 bromodomain looking down the acetyl-lysine binding pocket in which surface exposed conserved residues among bromodomains are indicated in purple

Structure of the chromodomain module a | Structure of chromodomain from MOD1 A hydrophobic pocket is present between the 1 helix and 2 strand and is proposed to be a substrate-binding site. b | Electrostatic surface representation of the MOD1 chromodomain module looking down the putative methyl-lysine substrate-binding pocket. MOD1, mouse modified protein 1

Приклади білків, що містять домени, що розпізнають модифікацію гістонів

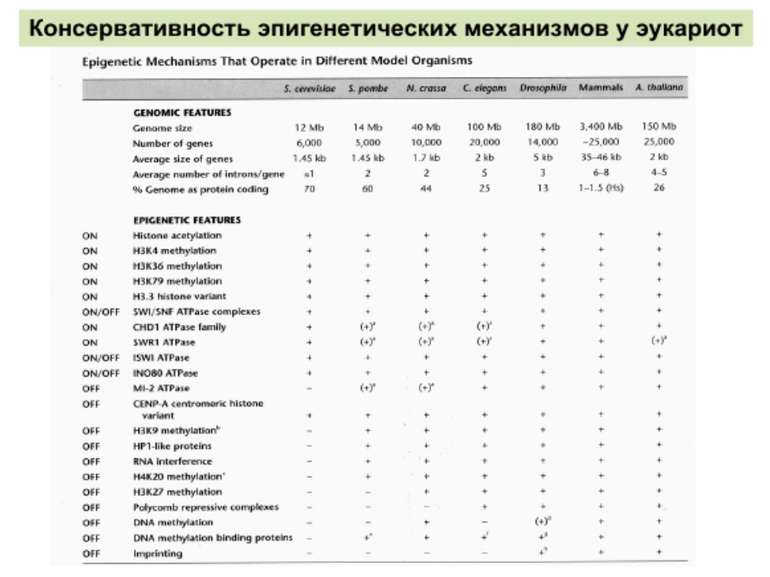
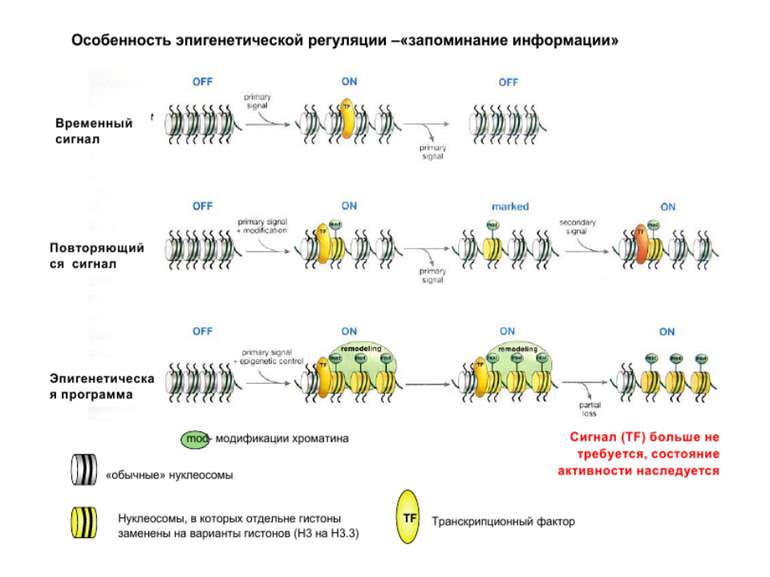
Як успадковується гістоновий код?

Як успадковується гістоновий код?

Як успадковується гістоновий код?

Взаємодія модифікацій гістонів
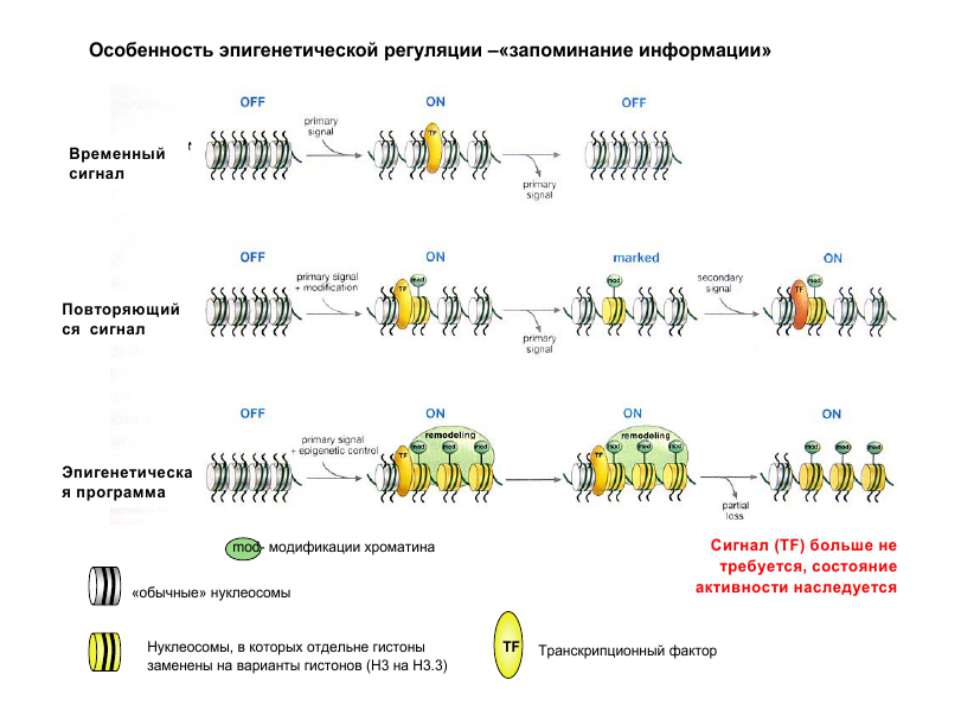

Схема рівнів активації хроматину, запропонована Kadonaga

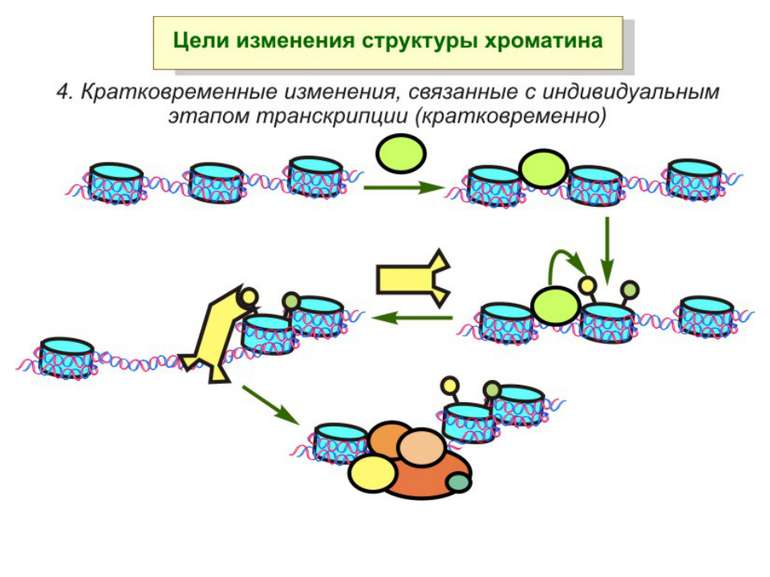
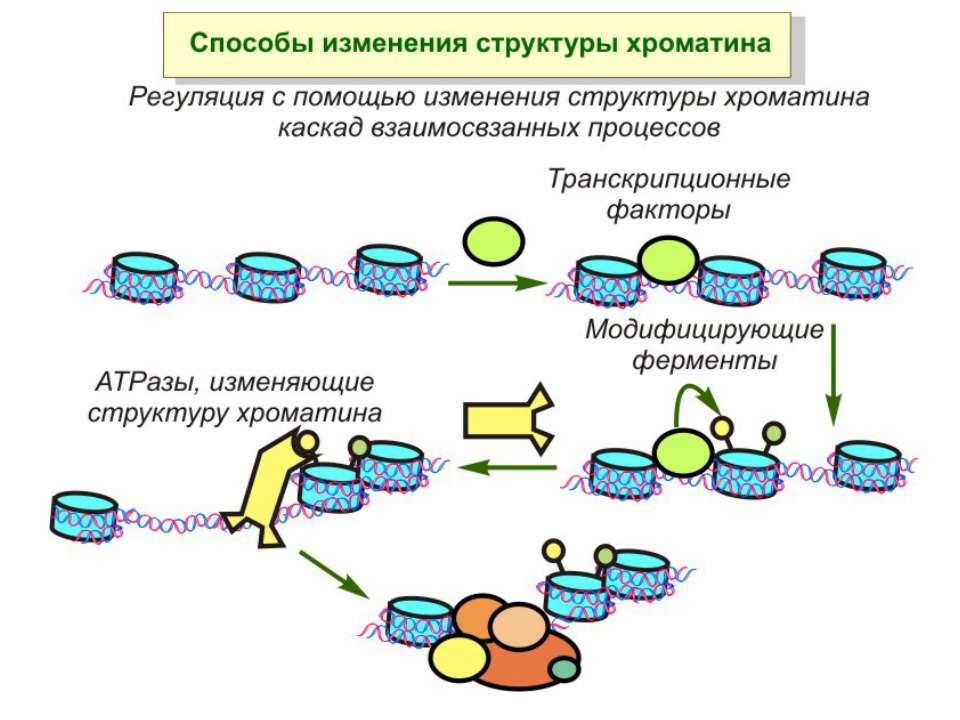
Після того, як відбулось ремоделювання структури хроматину HAC, SWI/SNF, ISWI комплексами, наступає час роботи РНК - полімерази II

Моделі транскрипції: На першому етапі із зарепресованим хроматином зв'язуються активатори, які сприяють роботі над усією послідовністю гена ремоделюючих комплексів, які, у свою чергу, роблять ДНК доступною для роботи РНК - полімерази II. Із зарепресованим хроматином зв'язуються активатори. Після цього уздовж послідовності гена проходить "первинна" РНК - полімераза, яка тягне на собі все необхідне для ремоделювання хроматину. При цьому полімераза транскрибує ген. Є експериментальні дані, що вказують на те, що первинний транскрипт відрізняється від наступних транскриптів з даного гена в даному активному стані. Слідом за "первинною" полімеразою починає працювати РНК - полімераза II, яка рухається вже по ремодельованому хроматину уздовж послідовності гена.

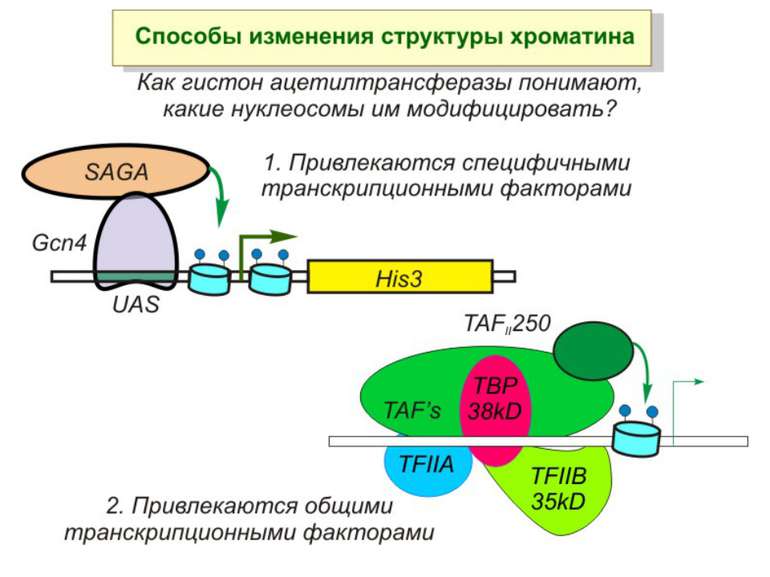
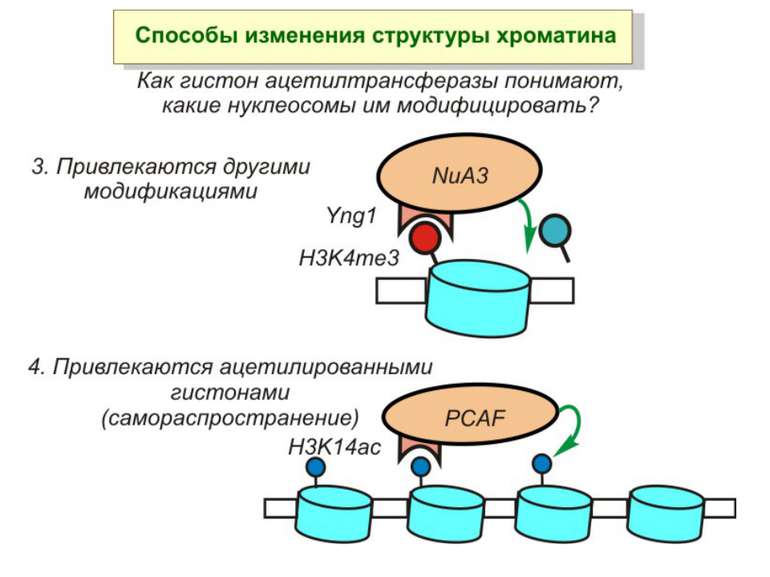
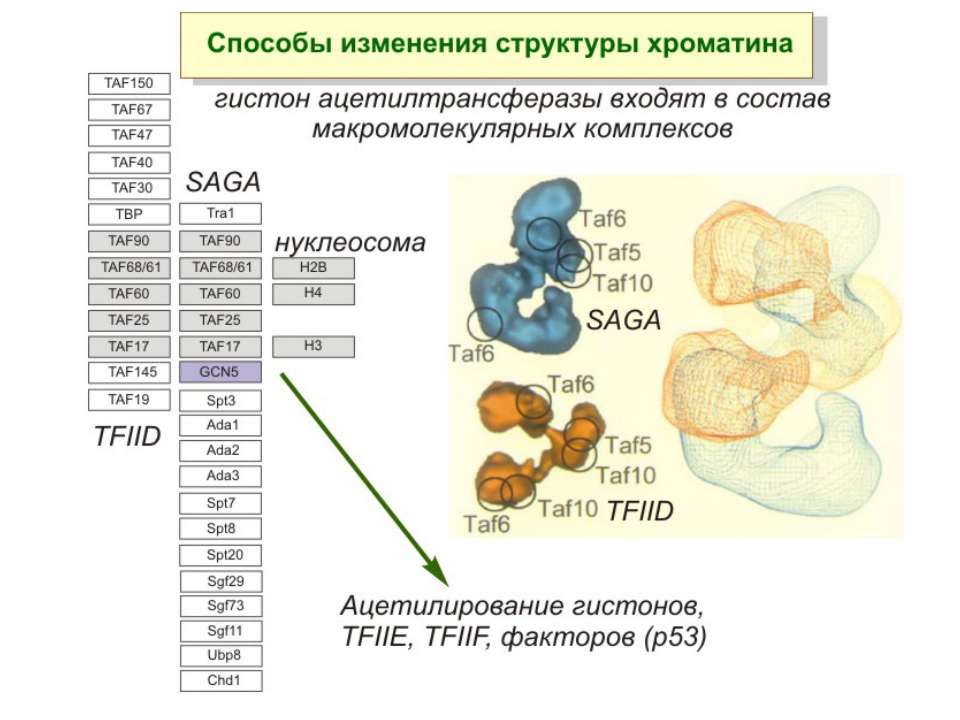
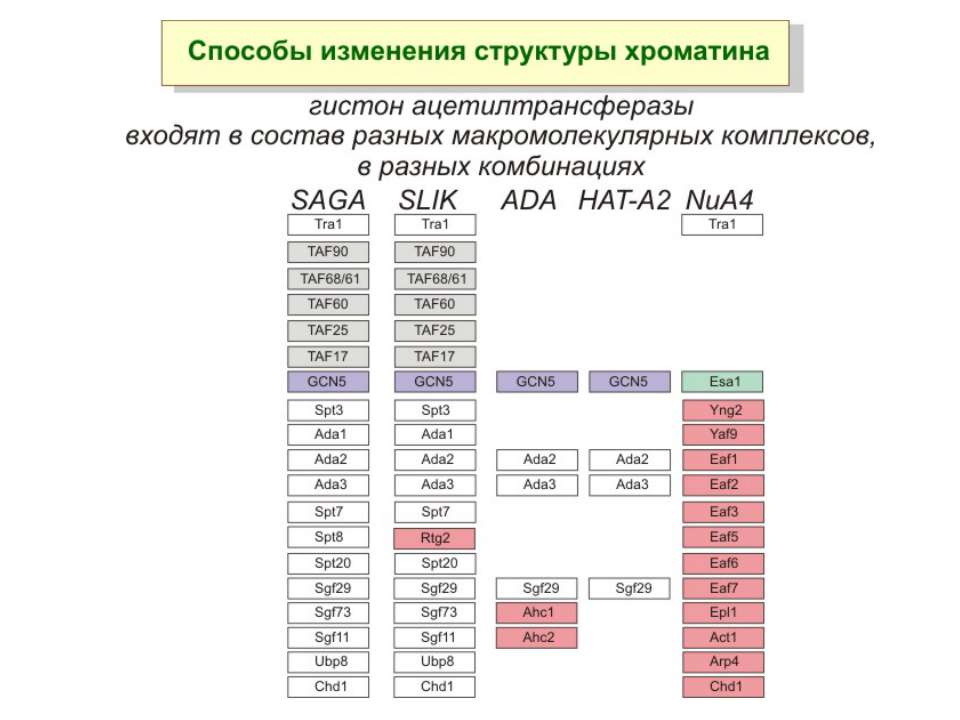
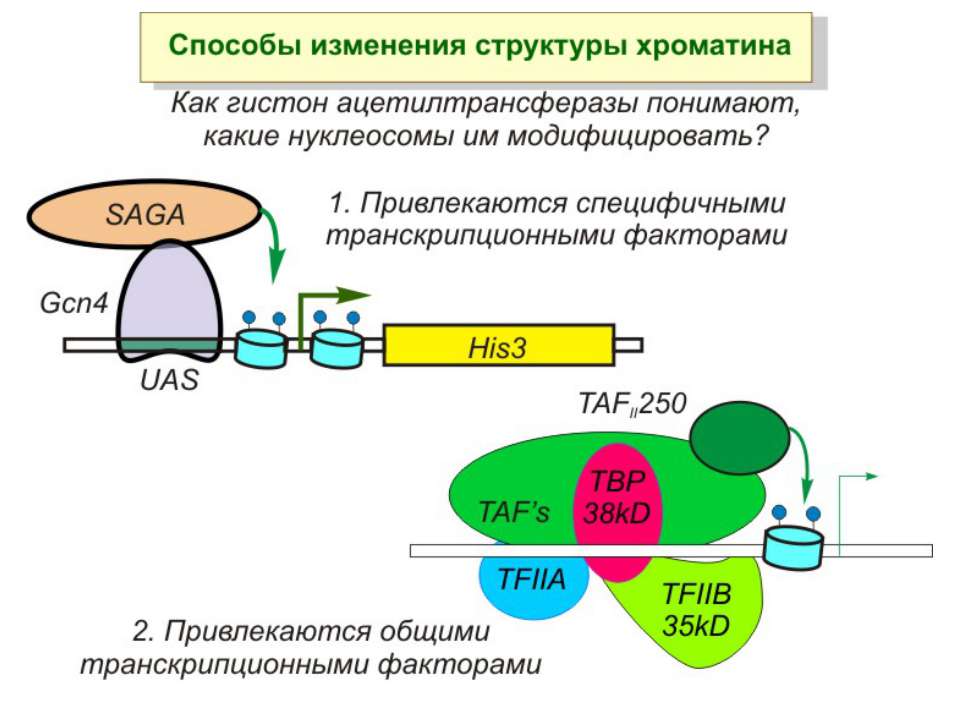
Приклади дотранскрипційних подій з дріжджовим промотором HO спочатку зв'язується SWI/SNF, потім SAGA. Після цих хроматин-модифікуючих комплексів зв'язується активатор, що «притягує» полімеразу за SRB/медіаторний комплекс ( що сидить на CTD домені РНК полімерази II). Викликана вірусом активація промотору інтерферону. Спочатку діє гістон-ацетилтрансфераза GCN5, потім гістон-ацетилтрансфераза CBP і відбувається посадка РНК-полімерази II. CBP приваблює SWI/SNF, який відсуває нуклеосому від TATA послідовності й, таким чином, звільняє місце для посадки TFIID. a 1-антитрипсиновий промотор. У неактивному стані з ним зв'язаний активатор HNF-1a, TFIIB і TBP. При активації спочатку збирається повний TFIID і приєднується пролімераза, потім – гістонацетилтрансферази CBP і PCAF. Після гіперацетилювання гістонів у районі промотору, приєднується HNF4 і SWI/SNF. Після цього відбувається ініціація.
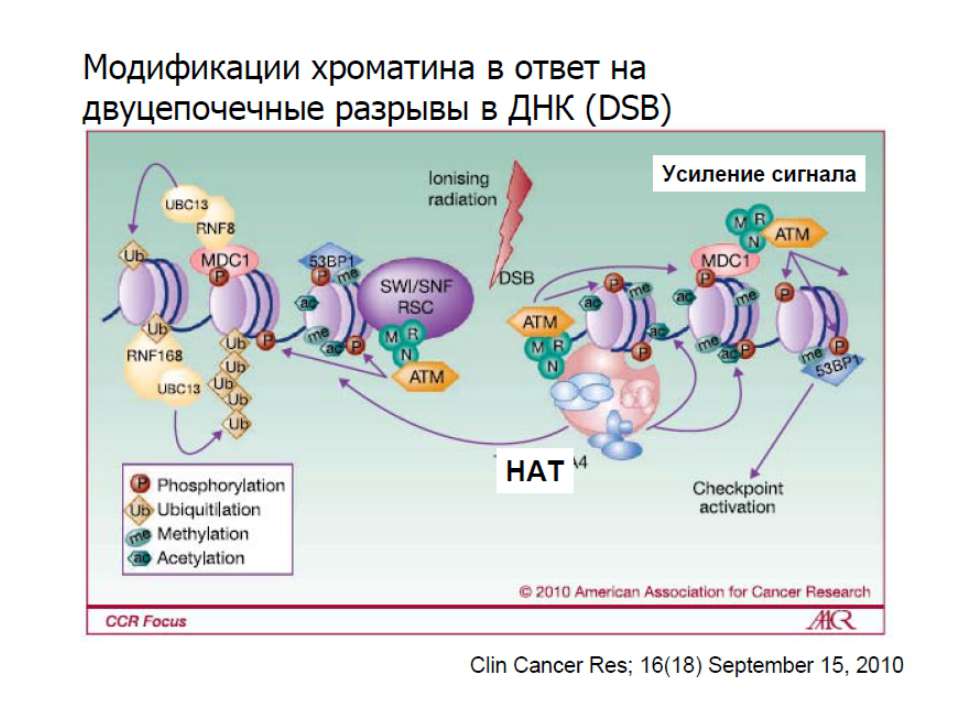
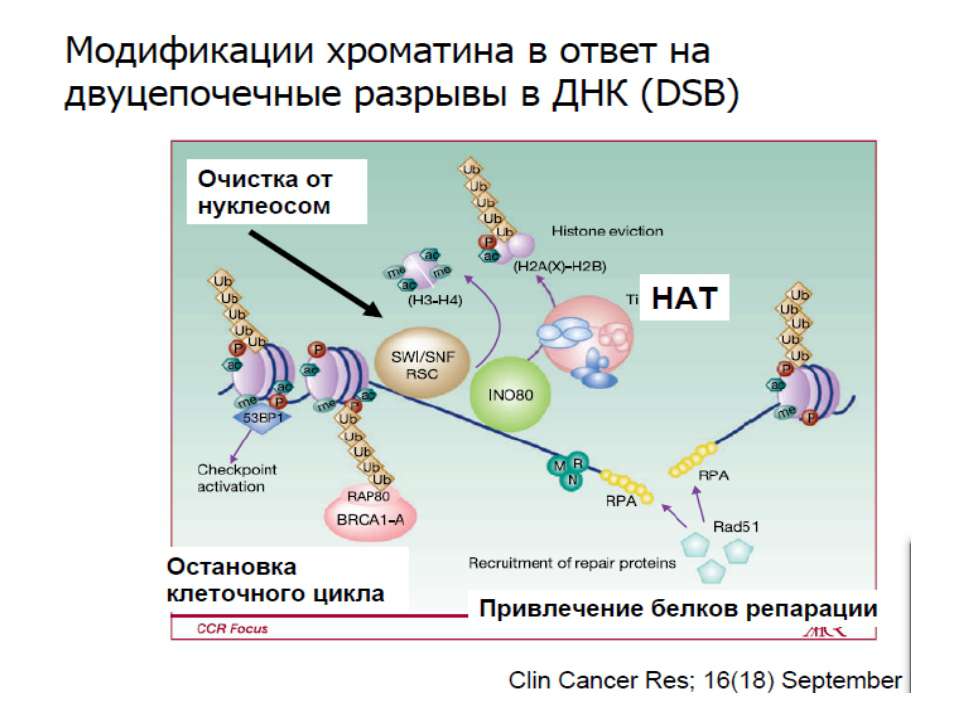

Транскрипційна регуляція еукаріот

Регуляція шляхом безпосередньої взаємодії з білками транскрипційного апарату

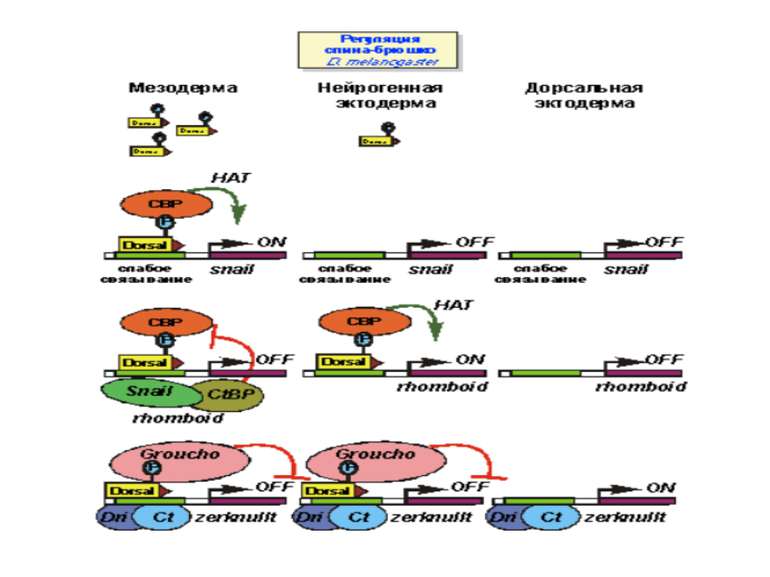
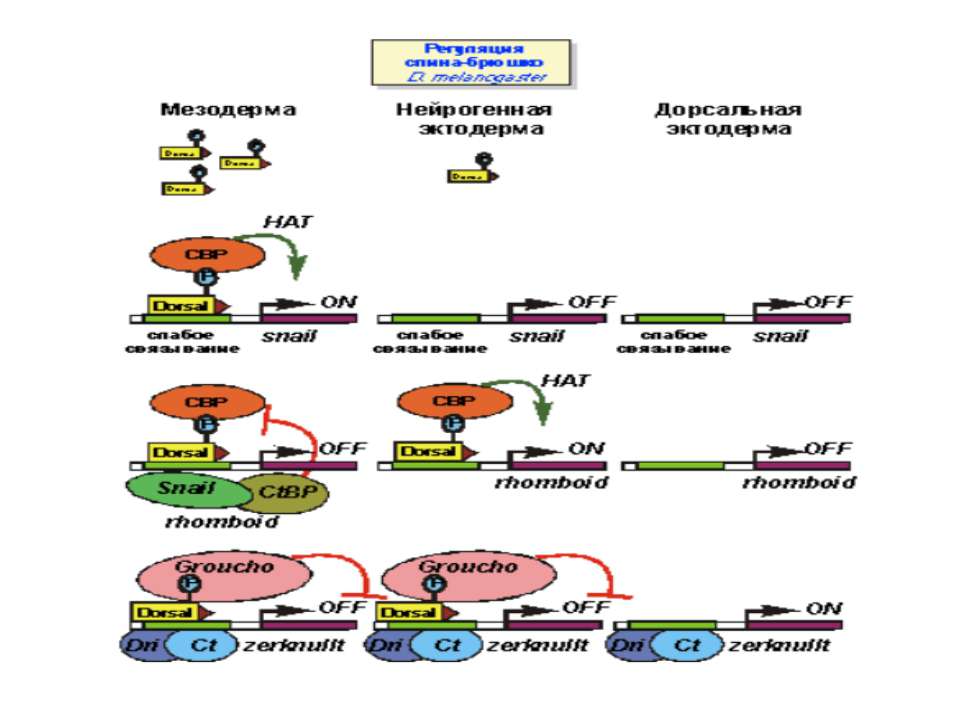
Регуляція диференціювання спинки та черевця у дрозофіли

Model for cooperation of the HAT, kinase and bromodomain modules for transcriptional activation In the first step, a histone H3 kinase is recruited to the promoter by the transcriptional activator to phosphorylate Ser10 on histone H3. b | A histone H4 HAT (histone acetyltransferase) complex is recruited to the promoter to acetylate a lysine residue on histone H4. c | The Gcn5 HAT complex is recruited to the promoter, possibly through multiple interactions including an interaction between the bromodomain of Gcn5 with acetyl-lysine modified histone H4 and an interaction between a phosphorylated histone H3 tail with the HAT domain. This recruitment results in the acetylation of Lys14 on histone H3. d | The effect of these post-translational histone modifications is to recruit RNA polymerase II and the GTFs (general transcription factors) to DNA for transcriptional activation. Ac, acetylation; P, phosphorylation.
Схожі презентації











Категорії











