Презентація на тему:
Розмноження на клітинному рівні
Завантажити презентацію






 S-фаз...")
 Складає 90% усього клітинного циклу Період найбільшої...")



 G1-фазова Cdk (Cdk4) S-фазова Cdk (Cdk2) M-фазова...")
 запускають под...")












 46 Мишадомашня 40 Кукурудза 20 ...")



")



 у рослинних клітинах Фл...")







 – 4 основні фази Цитокінез (поділ цито...")



 хромосом під час мітозу")







 Ро...")






 – розп...")





 кліт...")






...")
 клітини Приклади клітин: базальні епітеліальні к...")
 П...")
 клітини Приклади клітин: нейрони у центр...")









 Інтерфаза I – S-фаза (реплікація ДНК) відносно тр...")






 Поділ відповідає звичайному мітозу Ін...")






Розмноження на клітинному рівні
Завантажити презентаціюПрезентація по слайдам:

Розмноження на клітинному рівні Кафедра медичної біології ХНМУ-2010 Проф., д. мед. н. В.В. М’ясоєдов Ян Брейгель «Рай» Слайд-лекція

Питання лекції: Організація клітини у часі: життєвий цикл клітини і клітинний цикл Способи поділу клітини: мітоз, амітоз Мейоз – особливий спосіб поділу клітини Клонування

Організація клітини у часі: життєвий цикл клітини і клітинний цикл

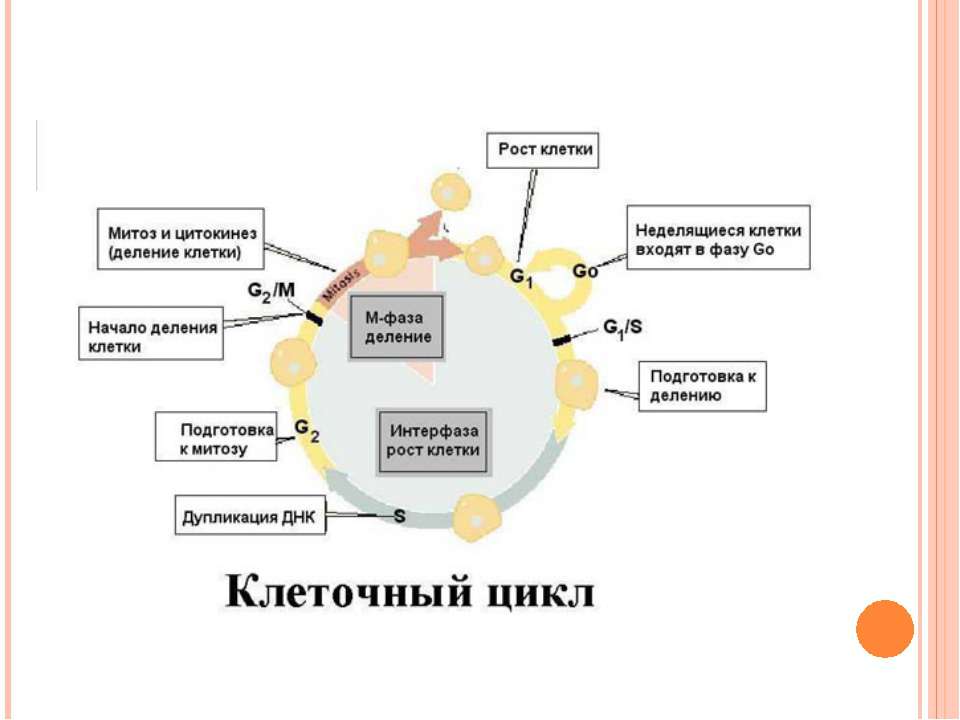
Життєвий і клітинний цикли У житті клітини розрізняють життєвий цикл і клітинний цикл Життєвий цикл – період від утворення клітини з материнської до наступного поділу або загибелі клітини Клітинний цикл включає підготовку до мітозу (інтерфазу) і мітоз. Друга назва процесу - мітотичний цикл.

Тривалість клітинного циклу в еукаріотичних клітинах – 10 – 20 годин Тривалість саме поділу – 1 година
 S-фаз...")
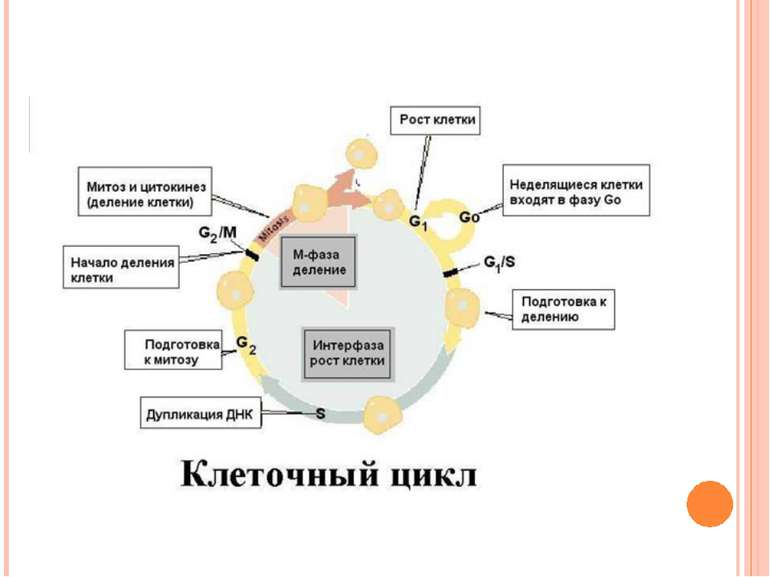
Фази клітинного циклу Інтерфаза G1-фаза – постмітотична (пресинтетична) S-фаза – синтетична G2-фаза – постсинтетична (премітотична) Мітоз Цитокінез G0 Точка рестрикції
 Складає 90% усього клітинного циклу Період найбільшої...")
Інтерфаза (G1 + S + G2) Складає 90% усього клітинного циклу Період найбільшої метаболічної активності Період підготовки до поділу Ядро інтактне, заповнене тонкими нитками - хромонемами

G1 – рост клітини, синтез РНК, білків, підготовка хромосом до поділу S – реплікація ДНК (і центросом) G2 – підготовка до мітозу, запасання енергії, синтез веретена поділу G0 Точка рестрикції

Життєвий цикл ентероцитів

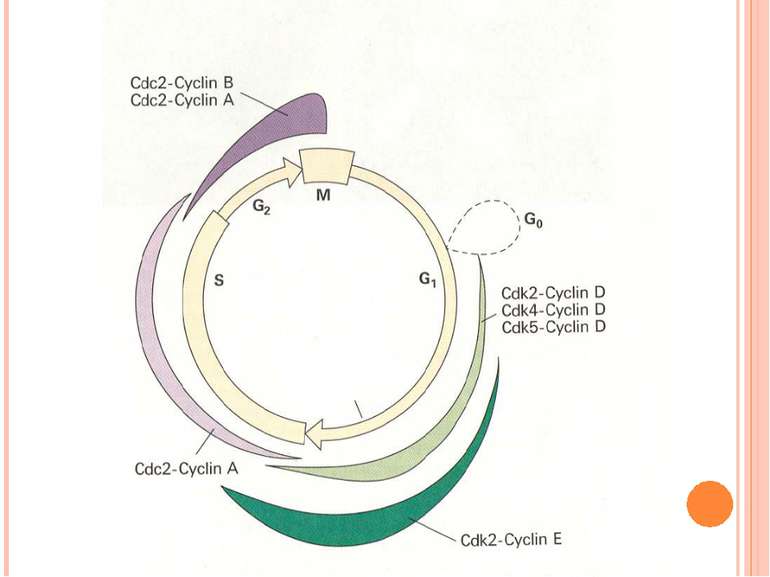
Контроль клітинного циклу Проходження клітини через клітинний цикл контролюється білками цитоплазми Головними серед білків у тваринній клітині є: Цикліни G1-циклін (циклін D) S- цикліни (цикліни E і A) М- цикліни (цикліни B і A) Рівень циклінів підвищується і знижується у залежності від фази циклу.

 G1-фазова Cdk (Cdk4) S-фазова Cdk (Cdk2) M-фазова...")
Циклін-залежні кінази (Cdk) G1-фазова Cdk (Cdk4) S-фазова Cdk (Cdk2) M-фазова Cdk (Cdk1) Рівні кіназ у клітині залишаються достатньо стабільними, але кожна з них повинна зв’язати відповідний циклін (рівень якого флюктуює) для активації Кінази приєднують фосфатні групи до білків, які контролюють клітинний цикл
 запускають под...")
Комплекс, що сприяє анафазі (АРС - anaphase-promoting complex) запускають події, що призводять до руйнування когезинів (що утримують сестринські хроматиди) і забезпечують розділення сестринських хроматид; руйнують мітотичні цикліни

Точки контролю клітинного циклу Контроль виходу з S-фази Контроль входу до мітозу Контроль переходу до анафази R – точка рестрикції S – точка старту Апоптоз Апоптоз Чи достатній об’єм клітини? Чи сприятливе середовище? Чи не ушкоджена ДНК? Чи реплікована ДНК повністю? Чи усі хромосоми пов’язані із веретеном? Ділитися або не ділитися? (взаємодія з ростовими факторами) Чи достатній розмір клітини? Чи не ушкоджена ДНК? Чи сприятливе середовище? Чи усі райони репліковані? G0

G0-фаза Клітина може виходити з клітинного циклу тимчасово або назавжди. Вона виходить із циклу в G1 і входить до стадії, що має назву G0. Частина G0–клітин екстремально диференційовані: вони ніколи не вступають повторно до клітинного циклу і виконують свої функції до загибелі (нейрони, кардіоміоцити). Інші G0–клітини можуть знову вступати до клітинного циклу. Більшість лімфоцитів у крові людини знаходяться у G0 стані. Але при сильній стимуляції антигенами вони можуть вступати до G1-фази і проходити S фазу і фазу мітозу. Ракові клітини не можуть переходити до G0–фази і постійно повторююють клітинний цикл. G0 Точка рестрикції

Хромосоми У еукаріот у періоді G1 хромосоми містять одну молекулу ДНК у вигляді 30 нм волокон, пов’язану з: великою кількістю гістонів; невеликою кількістю різних негістонових білків, більшість із яких – фактори транскрипції. Гистоні – это белки, богатіе лизином и аргинином, поэтому положительно заряженніе. По этой причине они прочно связіваются с отрицательно заряженніми фосфатами ДНК.

Від ДНК до метафазної хромосоми

Упаковка ДНК у хромосоми

Перед приготуванням клітини до поділу мітозом кожна хромосома подвоюється (у S фазу клітинного циклу) Із початком мітозу подвоєні хромосоми конденсуються, вони можуть бути пофарбовані й бути побаченими під світловим мікроскопом Подвоєні хромосоми називаються діадами.

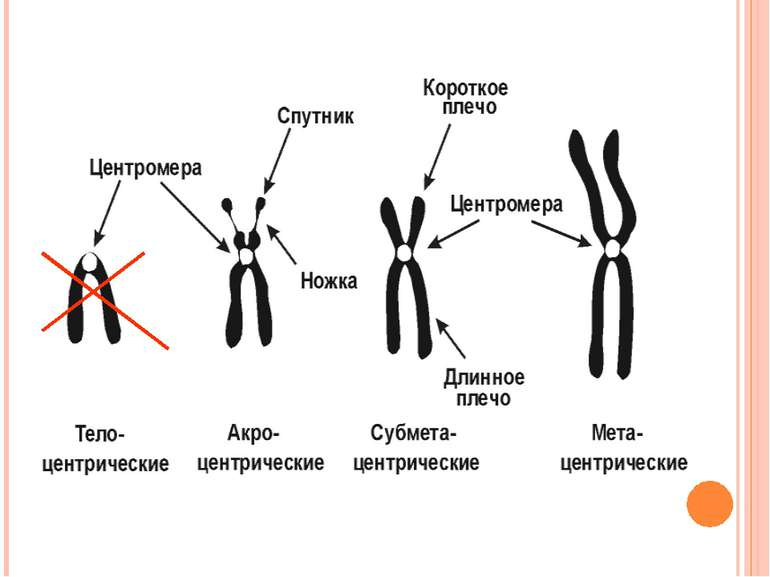
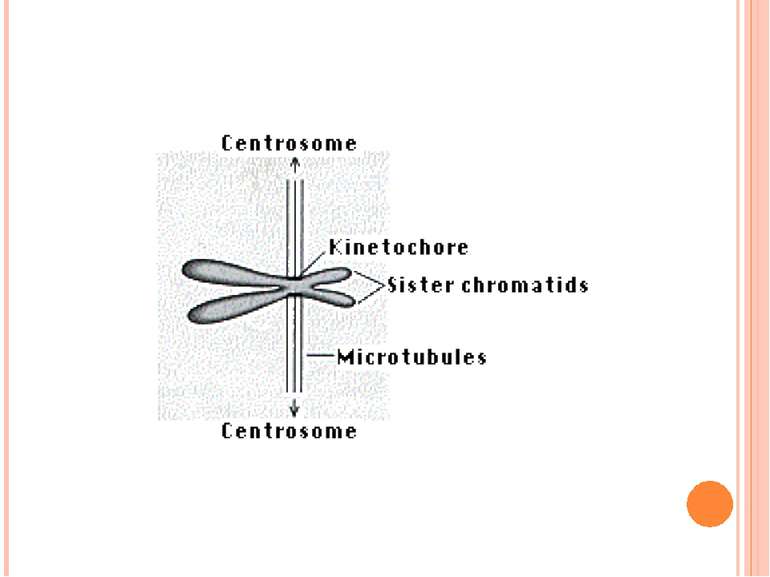
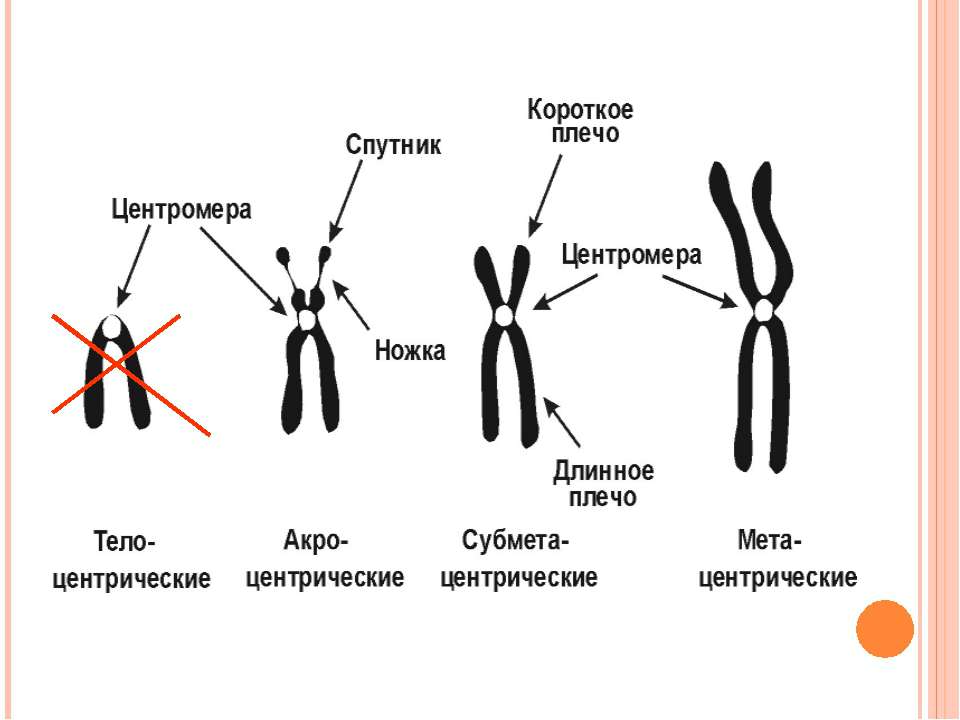
Подвоєні хромосоми утримуються разом в області центромери Частини подвоєних хромосом мають назву сестринських хроматид Кінетохор – це комплекс білків, що формуються в центромері й беруть участь у розділенні сестринських хроматид у анафазі мітозу Короткі плечі позначаються як p плечі; довгі - як q плечі. Забарвлення за допомогою барвника Гімза виявляє смуги, які називаються G смугами. G смуги нумеруються і використовуються в якості адреси генів Теломери Супутниковий район Вторинна перетинка Центромерний район (первинна перетинка) Теломери Сестринські хроматиди Коротке плече р Довге плече q

Метафазна хромосома Роль кінетохора у розділенні хроматид

Хромосома Мікротрубочки Кінетохори

Каріограма людини - схематичне зображення хромосом, розташованих за їхніми розмірами

Число хромосом Соматичні клітини містять дві копії геному і називаються диплоїдними (2n) Цей набір утворений гомологічними парами, кожний член яких походить із гамет кожного з батьків Гамети містять гаплоїдний набір (n) хромосом

Вміст ДНК у гаплоїдному наборі позначається с Число хромосом у геномі позначається n У людей с = 3,5 × 10-12 г n = 23 Вміст ДНК у диплоїдних клітинах – 2с, а число хромосом - 2n
 46 Мишадомашня 40 Кукурудза 20 ...")
Диплоїднінабориорганізмів Homosapiens(людина) 46 Мишадомашня 40 Кукурудза 20 Drosophilamelanogaster(плодовамушка) 8 Собакадомашній 78

Каріотип Каріотип – це повний набір хромосом у клітині організму Найчастіше за усе каріотип вивчається у метафазі мітозу, коли усі хромосоми представлені діадами

Каріотип жінки Каріотип жінки містить 23 пари гомологічних хромосом: 22 пари аутосом 1 пару X хромосом Бальдунг Грін Ганс Три віки і смерть

Каріотип чоловіка Каріотип чоловіка містить: 22 пари аутосом одну X хромосому одну Y хромосому Бара Едуард Корнуольський пейзаж із фігурами і олов’яними рудниками
")
Ідіограма людини, хворої на синдром Дауна (+21)

Вміст хромосомної ДНК людини під час клітинного циклу Хромосоми містять одну спіраль ДНК від анафази мітозу до подвоєння ДНК у S фазу. З цієї стадії до закінчення метафази мітозу хромосома містить дві хроматиди, кожна з яких містить молекулу ДНК, тобто на хромосому приходиться дві молекулі ДНК. Вміст ДНК диплоїдної клітини перед S фазою - 2с (подвоєний вміст ДНК гаплоїдної клітини), тоді як між S фазою і мітозом він дорівнює 4с. 2n2c 2n4c

Способи поділу клітини: мітоз, амітоз

Способи поділу клітини: мітоз і амітоз Основні способи поділу соматичних клітин – мітоз і амітоз мітоз (грец. мітос – нитка) – непрямий поділ клітини, переважний тип поділу соматичних клітин еукаріот Дочірні клітини, що утворюються при мітозі, генетично ідентичні материнській
 у рослинних клітинах Фл...")
поділ клітин уперше був описаний Страсбургером (1875) у рослинних клітинах Флемінгом (1879) у тваринних клітинах Термін «мітоз» було введено Флемінгом у 1880 році

Амітоз Амітоз – прямий поділ клітини, при якому генетичний матеріал не подвоюється (?) і розподіляється (рівномірно або нерівномірно) поміж дочірніми клітинами Характерний для деяких одноклітинних організмів Зустрічається у хрящевій, сполучній тканинах, у ракових клітинах Поділ амеб

При амітозі і клітина, і ядро подовжуються і розділяються посередині Дочірні клітини – приблизно рівні утворення Амітоз

У високоорганізованих організмів розрізняють амітоз двох типів: що призводить до утворення багатоядерних клітин (в епітелії, печінці), які далі не діляться, старіють і гинуть що призводить до розділення однієї клітини на дві (у хрящі, пухкій сполучній тканині) з утворенням ізогенних груп клітин із однієї материнської

Мітоз

Мітоз При поділі еукаріотичної клітини на дві, кожна дочірня клітина повинна отримати повний набір генів (для диплоїдних клітин 2n) пару центріолей (у тваринних клітинах) певну кількість мітохондрій певну кількість рибосом, частину ЕПР, та інших органел

Забезпечення дочірніх клітин точним диплоїдним набором генів потребує великої точності Фото дає графічне уявлення проблеми. Показано не більше 3% однієї молекули ДНК із хромосоми людини (після видалення гістонів). Розуміючи, що це тільки 3% ДНК лише однієї із 46 хромосом, можно уявити проблему, із якою зустрічається клітина перед поділом. Проблема вирішується за допомогою мітозу!

Фази мітозу
 – 4 основні фази Цитокінез (поділ цито...")
Мітоз включає: Каріокінез (поділ ядра) – 4 основні фази Цитокінез (поділ цитоплазми)

Профаза Дві центросоми клітини, що містять по парі центріолей, пересуваються до протилежних полюсів клітини. Формується мітотичне веретено. Воно утворене нитками, які містять ~20 мікротрубочок. Мікротрубочки утворюються з мономерів тубуліну в цитоплазмі і ростуть із кожної центросоми. Хромосоми стають більш короткими і компактними.

Прометафаза Ядерна оболонка розбирається у зв’язку із порушенням розчинності ламін, що стабілізують внутрішню мембрану. Білкова структура, кінетохор, знаходиться в центромері кожної хроматиди. Разом із руйнуванням ядерної оболонки нитки веретена приєднуються до кінетохору. У кожній діаді один кінетохор приєднує одну хроматиду до одного полюсу, а другий - іншу сестринську хроматиду до іншого полюсу. Порушення приєднання нитки веретена до кінетохору перериває процес.
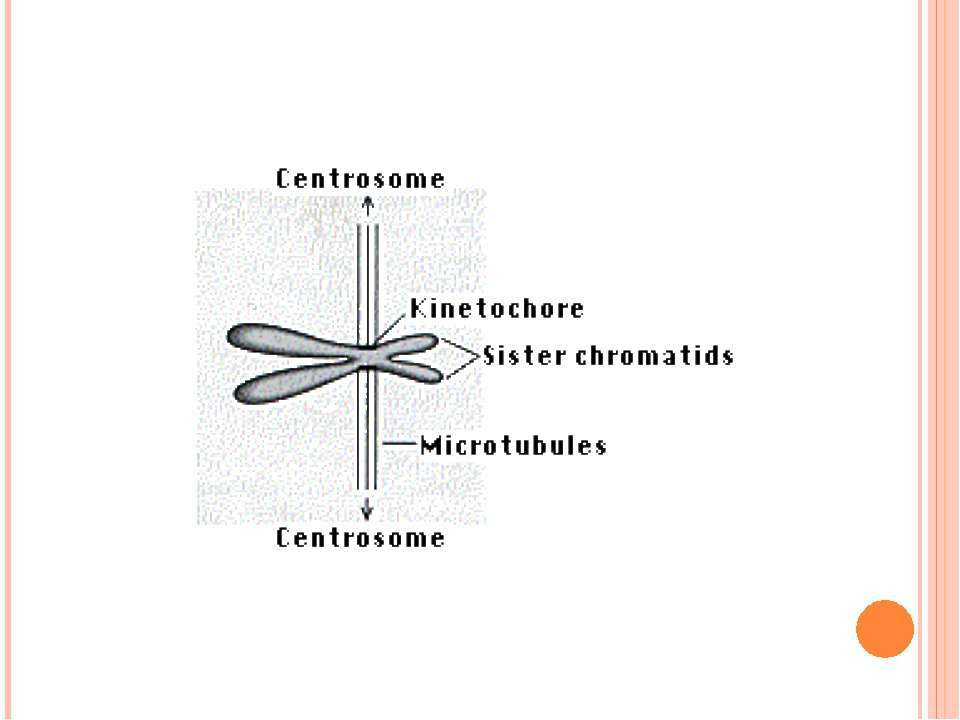

У метафазі усі діади займають однакове положення в екваторі клітини і утворюють метафазну пластинку. У цей час хромосоми найбільш компактні. У анафазу сестринські кінетохори раптово розділяються і кожний пересувається до відповідного полюсу, тягнучи за собою одну хроматиду. Розподіл сестринських хромосом залежить від руйнування когезинів, які утримують їх разом.
 хромосом під час мітозу")
Розподіл (сегрегація) хромосом під час мітозу

Кінетохор і веретено поділу

Мітози у зародку дрозофіли

Телофаза Навколо кожної групи хромосом формується ядерна оболонка, хромосоми деконденсуються починається поділ цитоплазми

Цитокінез За мітозом зазвичай іде поділ клітини. Однак, є випадки (у ембріонів комах), коли хромосоми підлягають мітозу без поділу клітини. Тому, є спеціальний термін – цитокінез – для позначення поділу клітини на дві. У тваринних клітинах актинові філаменти формують борозну по периметру клітини. Поглиблення борозни призводить до перетяжки клітини на дві. У рослинних клітинах формується клітинна пластинка, яка синтезується комплексом Гольджі.

200 мкм 25 мкм Цитокінез тваринної клітини відбувається завдяки актин-міозиновому кільцю Борозна поділу між двома бластомерами жаби

Скорочувальне кільце

Мітоз
 Ро...")
Значення мітозу Розмноження – наприклад у одноклітинних організмів (амеба) Розвиток, рост і генетична сталість – у багатоклітинних мітоз – це частина ембріонального розвитку, росту, регенерації і спадковості Клітинний метаболізм

Мітоз без цитокінезу Мітоз без цитокінезу утворює масу цитоплазми із багатьма ядрами. Приклад: стадія вільних ядер при ембріональному розвитку мух, подібних до Drosophila

Ендореплікація Ендореплікація – це реплікація ДНК під час S фази клітинного циклу без наступного мітозу та/або цитокінезу Ендореплікація відбувається у визначених клітинах тварин і рослин

Варіанти ендореплікації: реплікація ДНК із повним мітозом, але без цитокінезу (+ М, ─ цитокінез). повторна реплікація ДНК без формування нових ядер у телофазі (+++реплікація, ─ ядра у телофазі). Результатом може бути: Поліплоїдія: репліковані хромосоми залишаються в клітині Політенія: репліковані хромосоми залишаються в лінії, формуючи гігантські хромосоми. різноманітні проміжні стани між 1 та 2

Поліплоїдія У поліплоїдних клітинах число хромосом на n більше, ніж у диплоїдній клітині (2n ): триплоїдна (3n), тетраплоїдна (4n) ... Поліплоїдія зазвичай обмежена визначеними клітинами у тварин, такими як: гепатоцити; мегакаріоцити; мегакаріоцити, з яких утворюються тромбоцити, можуть проходити через 7 S фаз, утворюючи гігантські клітини з одним ядром, що містить 128n хромосом. Їхня фрагментація дає тромбоцити. гігантські трофобластні клітини у плаценті. Поліплоїдія у рослин – дуже часте явище

Поліплоїдія у тварин Поліплоїдія у тварин дуже нечаста. Вона виявлена у деяких комах, риб, амфібій і рептилій. До недавнього часу про поліплоїдію у ссавців не було відомо. Проте, 23 вересня 1999 року у журналі Nature було повідомлення про поліплоїдного щура (тетраплоїд; 4n = 102), знайденого в Аргентині Поліплоїдні клітини більші, ніж диплоїдні; в ядрах клітин збільшена кількість ДНК. Клітини печінки Аргентинського щура більші ніж клітини диплоїдів, а його сперматозоїди порівняно величезні. Голівка нормального спермія ссавця містить біля 3.3 пікограмів (10-12 g) ДНК; спермії щура містить 9.2 пг.

Політенія Найбільш вивченими прикладами політенії є гігантські хромосоми, знайдені у мух. Мікрофотографія показує політенні хромосоми клітин слинних залоз Drosophila melanogaster. Такі хромосоми також виявляються в інших великих, активних клітинах. Кожні 4 пари хромосом дрозофіли (каріотип) проходять 10 циклів реплікації ДНК. Материнські і батьківські гомологи – як і усі їхні дуплікати - випрямлені у точній відповідності один із одним Тому кожна хромосома уявляє собою кабель, який містить 2048 ідентичних ланцюгів ДНК. Вони такі великі, що можуть бути побаченими в інтерфазі навіть у слабкому світловому мікроскопі Функція політенії – ампліфікація генів, посилення експресії генів
 – розп...")
Політенні хромосоми мають регіони, які називаються «пуфами» (пуховики) – розпушені регіони Картина пуфів різноманітна у різноманітних типах клітин і змінюється із зміною стану клітини Пуфи уявляють собою регіони інтенсивної транскрипції генів

Порушення мітозу, соматичні мутації Наслідком порушення мітозу (патологічного мітозу) є дочірні клітини із різними каріотипами Патологічний мітоз – одна із причин соматичної анеуплоїдії ( -1, +1, -2, +2 …) Патологічний мітоз спостерігається при: променевій хворобі вірусних інфекціях раку

При порушенні мітозу можуть утворюватися: хромосомні мостики мікроядра пошкодження центромер склеювання хромосом та ін.

Патологічні мітози в клітинах пухлини

Пікнотичні ядра

Кільцева хромосома
 кліт...")
Соматичні мутації Це мутації, які відбуваються в соматичних (нестатевих) клітинах Властивості соматичних мутацій: Обмежені одним організмом Чим раніше в онтогенезі мутація, тим у більшій кількості клітин вона проявляється Фенотипові прояви мутації залежать від кількості клітин із мутацією Не передаються наступному поколінню, але можуть знижувати репродуктивний потенціал

Рост клітини, фактори росту Фактори росту – це фактори, які забезпечують виживання і проліферацію клітин Факторами росту можуть бути білки, пептиди, стероїди

Поліпептидні фактори росту поділяються на декілька суперродин: суперродина інсуліноподібних факторів росту (інсулін, релаксин та ін.) суперродина епідермальних факторів росту суперродина бомбезину (бомбезин, літорин, нейротензин) суперродина факторів росту фібробластів суперродина трансформуючих факторів росту суперродина факторів росту тромбоцитів цитокіни

Поліпептидні фактори росту пов’язуються із специфічними рецепторами плазматичної мембрани і викликають відповідну реакцію клітини Фактор росту Рецептор

Злоякісна трансформація клітин Зміни регуляції проліферації клітин можуть викликати злоякісну трансформацію клітин клітини, здатні реагувати на зовнішні фактори росту, починають секретувати їх самі клітини, які продукують фактори росту і не мають рецепторів до них, починають продукувати рецептори У клітинах навіть за відсутності дії на них факторів росту запускаються механізми синтезу ДНК і мітозу

Властивості злоякісних клітин Невпинна здатність до поділу внаслідок аномальної реакції на сигнали контролю поділу (відсутність контактного гальмування) Потребують менше факторів росту, ніж нормальні клітини Здатність ділитися багаторазово, не знижуючи мітотичного потенціалу Здатність проростати в інші тканини і стимулювати рост капілярів для живлення

Мітотична активність тканин Показник мітотичної активності тканин – це число клітин, що діляться мітозом, на 1000 вивчених клітин гістологічного препарата Для вивчення мітотичної активності використовується колхіцин (припинення мітозу), визначення включення міченого тимідину в нові молекули ДНК
...")
Клітини за мітотичною активністю поділяються на: Мітотично активні (лабільні) Зворотньо постмітотичні або «ті, що покояться» (відносно стабільні) Незворотньо постмітотичні (постійні)
 клітини Приклади клітин: базальні епітеліальні к...")
Мітотично активні (лабільні) клітини Приклади клітин: базальні епітеліальні камбіальні клітини усіх типів епітелію і гемопоетичні стовбурові клітини у кістковому мозку. Діляться протягом усього життя, є джерелом для відновлення клітин, які безперервно гинуть. Мають короткий G0 період. Зрілі диференційовані клітини у цих специфічних тканинах не можуть ділитися; їхня кількість підтримується поділом їхніх стовбурових лабільних клітин.
 П...")
Зворотньо постмітотичні або “ті, що покояться” клітини (відносно стабільні) Приклади клітин: паренхіматозні клітини найбільш важливих залозистих органів (печінка, підшлункова залоза) і мезенхімальні клітини (фібробласти, ендотеліальні клітини). Клітини мають тривалий термін існування і тому характеризуються низькою мітотичною активністю. Вони залишаються у фазі G0 протягом тривалого часу (часто роками), але зберігають здатність до поділу, коли входять до мітотичного циклу за потребою.
 клітини Приклади клітин: нейрони у центр...")
Постійно (незворотньо постмітотичні) клітини Приклади клітин: нейрони у центральній і периферичній нервовій системі і клітини міокарда. Постійні клітини не мають будь-якої здатності до мітотичного поділу в постнатальному житті. Пошкодження постійних клітин завжди супроводжується формуванням рубця. Повна регенерація неможлива. Втрата постійних клітин є незворотною і, якщо некроз великий за обсягом, це може призводити до порушення функції органів.

Мейоз – особливий спосіб поділу клітини

Мейоз, його біологічне значення Кожний організм є смертним. Необхідним є розмноження! Деякі найпростіші і більшість багатоклітинних зберігають свої види статевим розмноженням (об’єднання двох гамет → зигота → тканини і органи) Диплоїдність гамет призводила б до появи нежиттєздатних поколінь Гамети повинні бути гаплоїдними (n)!

Мейоз – спеціальна форма поділу генеративних клітин, яка призводить до утворення гаплоїдних гамет (сперматозоїдів і яйцеклітин у людини)

Визначений набір диплоїдних клітин організма утворює гермінальну лінію (клітини зародкового шляху), що бере участь у розмноженні Вони дають початок спеціалізованим диплоїдним клітинам у яєчниках і сім’яниках, які можуть поділятися мейозом і призводить до утворення гаплоїдних гамет (сперматозоїдів і яйцеклітин)

Життя людей іх хромосомної точки зору Гаплоїдні сперматозоїди і яйцеклітини утворюються внаслідок мейозу диплоїдних клітин-попередниць. У заплідненій яйцеклітині хромосоми сперматозоїда і яйцеклітини розділені, коли знаходяться у чоловічому та жіночому пронуклеусах. Вони об’єднуються під час першого мітозу.

Первинні гермінальні клітини мігрують у гонади ембріона і підлягає декільком мітотичним поділам (у чоловіків значно більше, ніж у жінок, що може бути фактором, який пояснює статеві відміни у частоті мутацій) з утворенням овогоніїв у жінок і сперматогоніїв у чоловіків. Подальший рост і диференціація призводять до утворення первинних овоцитів у яєчниках і первинниіх сперматоцитів у яєчках. Ці спеціалізовані клітини можуть підлягати мейозу

Фази мейозу Мейоз включає два послідовних клітинних поділи (мейоз I і мейоз II), але лише один процес реплікації ДНК, тому продукти є гаплоїдними

Порівняння мітозу й мейозу

Мейоз
 Інтерфаза I – S-фаза (реплікація ДНК) відносно тр...")
Мейоз I (редукційний поділ) Інтерфаза I – S-фаза (реплікація ДНК) відносно триваліша, ніж в інтерфазі мітозу, а G2-фаза більш коротка або відсутня Профаза I – дуже тривалий і складний процес, поділяється на 5 субфаз

Профаза I

«Лампові щітки» у диплотені Пара хромосом Дві хроматиди одного гомолога

Мікрофотографія бівалента хромосом типа «лампові щітки» Стрілками показано хіазми

Метафаза I – біваленти в метафазній пластинці (сестринські хроматиди гомологів поєднані з одним полюсом) Анафаза I – гомологічні хромосоми кожного бівалента мігрують до різних полюсів. У кожного полюса збирається гаплоїдний набір хромосом Телофаза I – рідко завершується до початку другого мітозу. Лише у деяких випадках ядерна оболонка може утворюватися навколо гаплоїдної групи хромосом. У більшості випадків мейоз II стартує без цих змін

Розподіл хромосом у мейозі

Другий мейотичний поділ є ідентичним мітозу, але перший поділ має важливі відміни, метою яких є генерування генетичної відміни між дочірніми клітинами. Це здійснюється двома механізмами: незалежним розподілом батьківських і материнських гомологів і рекомбінацією (кросинговером) Незалежний розподіл батьківських і материнських хромосом у мейозі I
 Поділ відповідає звичайному мітозу Ін...")
Мейоз II (екваційний, мітотичний поділ) Поділ відповідає звичайному мітозу Інтерфаза II – короткий період, реплікації ДНК немає (краща назва інтеркінез) Профаза II Метафаза II Анафаза II Телофаза II

У чоловіків внаслідок мейозу утворюється 4 сперматозоїда, а у жінок 1 яйцеклітина і 3 полярних тільця

Значення мейозу Зменшення числа хромосом. Гаплоїдність – найважливіше підґрунтя статевого розмноження Спадковість і мінливість. У зиготі й у всіх клітинах тіла в парі гомологічних хромосом – 1 батьківська і 1 материнська. Генетичне розмаїття людей – результат 3 процесів: випадкового розходження батьківських і материнських хромосом у мейозі I кросинговера у профазі мейоза I випадковості у об’єднанні гамет

Порівняння мітозу і мейозу Мітоз Мейоз Локалізація Усітканини Лишеуяєчкахіяєчниках Продукти Диплоїднісоматичніклітини Гаплоїднісперматозоїдиіяйцеклітини РеплікаціяДНКіподілклітин Унорміоднареплікаціяна одинподіл Лишеоднареплікація,аледваклітиннихподіли Тривалістьпрофази Коротка (~30хвуклітинахлюдини) МейозIтривалийіскладний;можетриватироками дозавершення Спарюваннягомологів Немає Є (умейозіI) Рекомбінація Рідкоабоувипадкаханомалії Унормі, поменшіймірі, одноразово у кожномуплечіхромосоми Дочірніклітини Генетичноідентичні Різноманітні(рекомбінаціяінезалежнийрозподілгомологів)

Клонування

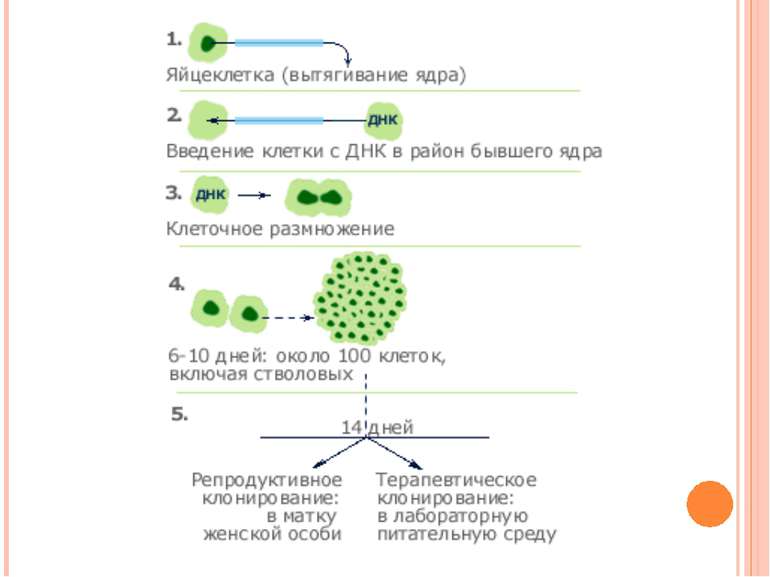
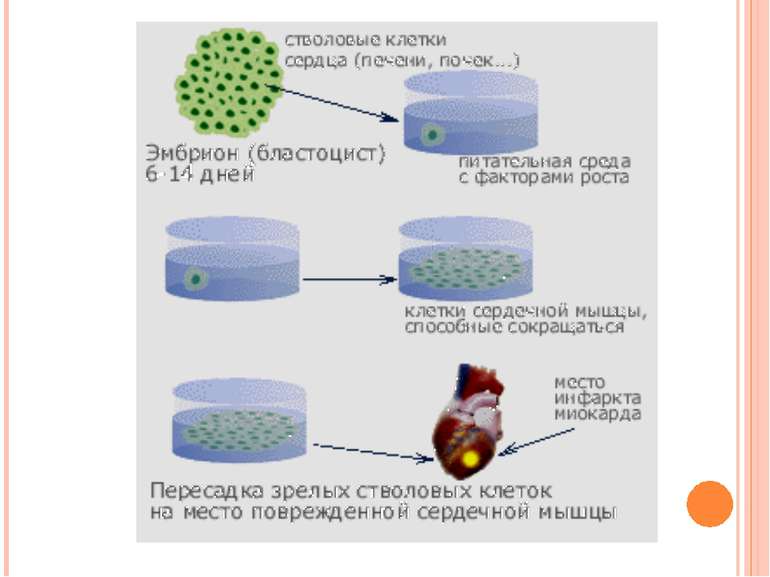
Клонування Клонування – це створення генетично ідентичних нащадків за допомогою нестатевого розмноження На початку 60-х років ХХ ст. було розроблено методи, що дозволяли успішно клонувати деякі вищі рослини і тварини
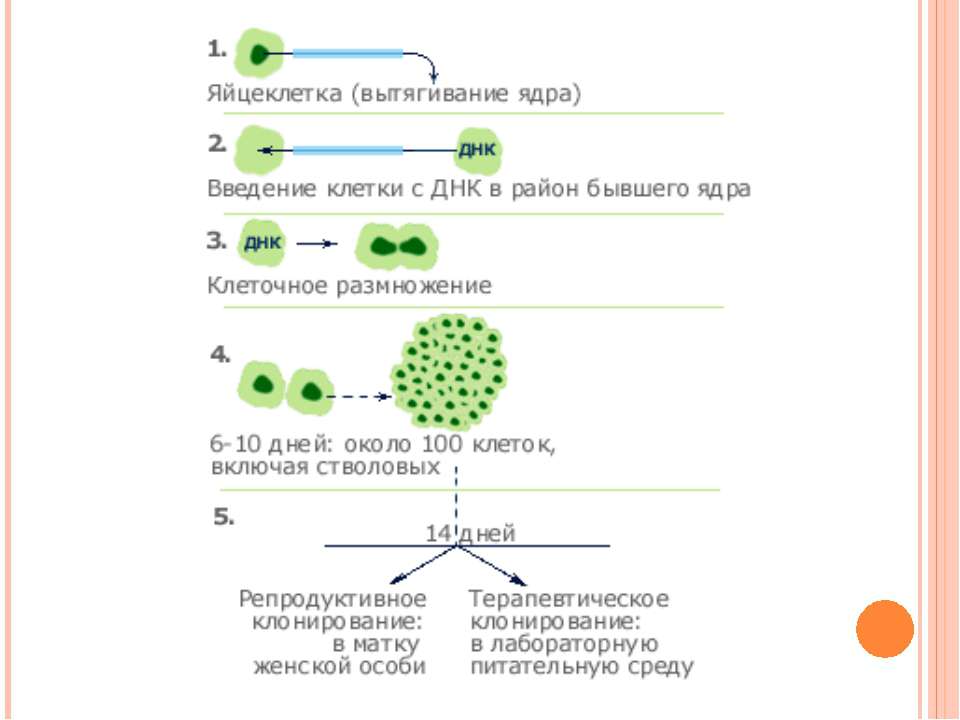

Репродуктивне клонування Клоновані людські ембріони
Схожі презентації











Категорії











