Презентація на тему:
Політика “умиротворення” агресора та її крах
Завантажити презентацію











")







 період може бути прямим і непрямим. За прям...")


Політика “умиротворення” агресора та її крах
Завантажити презентаціюПрезентація по слайдам:

Презентація з теми: Індивідуальний розвиток організму Підготував: Учень 10 класу Піщанської ЗОШ І-ІІІ ступенів Ім. Л.М. Дудки Плех Владислав

Індивідуальний розвиток організму від його зародження до смерті називається онтогенезом. Цей процес формування живого організму в кожного виду має свої особливості і тривалість, які склалися в процесі еволюції і є продуктом філогенезу. Онтогенез відбувається на базі єдності спадковості (внутрішні фактори), що історично склалася, і зовнішнього середовища (зовнішні фактори). В одноклітинних онтогенез збігається з клітинним циклом. У них він починається від поділу материнської клітини на дочірні й триває до наступного поділу утвореної клітини. У багатоклітинних організмів за безстатевого розмноження індивідуальний розвиток починається з однієї клітини або групи клітин, які дають початок безстатевому зачатку, і закінчується смертю. Початком онтогенезу за статевого розмноження вважають формування статевих клітин (овоґенез і сперматогенез). Це так званий проембріопальний (передзародковий) період розвитку. За ним іде ембріональний (зародковий, ембріогенез), який відбувається в зародкових або яйцевих оболонках, насінині. Він завершується народженням (вилупленням, проростанням), після чого починається постембріональний (післязародковий) період, що закінчується смертю організму. Особливості онтогенезу тварин. Онтогенез означає реалізацію генетичної програми. Зміна активності генів лежить в основі диференціювання тих чи інших клітин у необхідному напрямку на певних стадіях онтогенезу. У процесі ембріонального розвитку з'являються спочатку загальні ознаки типу, а потім послідовно — класу, ряду і, зрештою, виду. Це означає, що ембріони різних тварин з одним планом будови більш подібні між собою, ніж дорослі особини. Саме з цього Геккель вивів «біогенетичний закон». Етапи індивідуального розвитку організмів

Цей період триває від моменту запліднення яйцеклітини до початку вільного самостійного життя з власним пересуванням і живленням. В ембріональному розвитку людини вирізняються три періоди: початковий, зародковий і плідний. Початковий період охоплює перший тиждень ембріонального розвитку. Він починається з моменту запліднення і триває до початку імплантації зародка в слизову оболонку матки. В цей період розпочинається стадія дроблення. Зародковий період охоплює час від початку другого тижня після запліднення до кінця восьмого тижня, тобто від імплантації до завершення процесу органогенезу. Останній, плідний, або фетальний, період ембріонального розвитку людини починається з дев'ятого тижня після запліднення і триває до народження. В ембріональний період послідовно відбуваються процеси дроблення, гаструляції, органогенезу та гістогенезу. Зародковий (ембріональний) період розвитку Ембріон людини

Відразу після запліднення починається процес поділу зиготи, який називають дробленням, а клітини, які при цьому утворюються, — бластомерами. Перша і друга борозни дроблення проходять меридіонально, а третя — в екваторіальному напрямку. З кожним поділом кількість клітин збільшується, самі бластомери зменшуються вдвоє без зменшення ядра, зародок не росте. В подальшому співвідношення ядра і цитоплазми відновлюється. Характер дроблення залежить від кількості й розподілу жовтка в яйцеклітині. Якщо жовтка в клітині мало, середня кількість або зовсім немає, в процес дроблення втягується вся клітина, тобто відбувається повне дроблення (ізолецитальні яйцеклітини у ланцетника). У деяких видів тварин дробиться тільки невелика вільна від жовтка частина зиготи (зародковий диск), де міститься ядро. Таке дроблення називається неповним, частковим — у плазунів, птахів, першозвірів. Повне дроблення може бути рівномірним і нерівномірним. Повне нерівномірне дроблення властиве телолецитальним яйцеклітинам (амфібії). Завершується дроблення утворенням бластули, яка має вигляд порожнистої кулі з порожниною (бластоцель), що заповнена рідиною. Стінка бластули (бластодерма) складається з одного шару бластомерів. Якщо під час дроблення утворюється кулястий зародок без порожнини всередині, подібний до супліддя шовковиці, його називають морулою. Така бластула властива, наприклад, плацентарним ссавцям. У бластулі є дах (анімальний полюс із дрібнішими бластомерами) і дно (вегетативний полюс, бластомери більші). Дроблення та утворення бластули (на прикладі ссавців) Дроблення ізолецитальної яйцеклітини ланцетника та утворення бластули: а — зигота (1 — полярне тільце, 2 — анімальний полюс, 3 — вегетативний полюс); б — стадія двох бластомерів; в — стадія чотирьох бластомерів; г — стадія восьми бластомерів; д, е — стадії 16 і 32 бластомерів — морула; є — бластула (4 — дах бластули, 5 — дно бластули, 6 — бластоцель)

Типи дроблення яйцеклітин: а — повне рівномірне ізолецитальної яйцеклітини; б — повне нерівномірне телолецитальної яйцеклітини; в — дискоїдальне (часткове) телолецитальної яйцеклітини; г — поверхневе (часткове) центро-лецитальної яйцеклітини

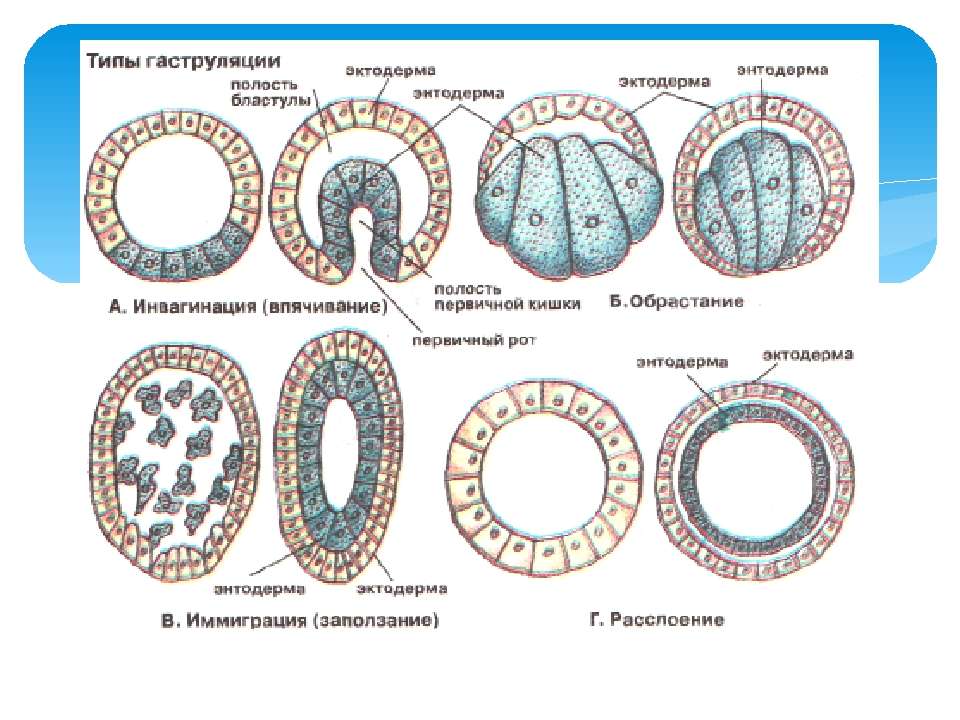
Сукупність процесів, які забезпечують утворення гаструли, називають гаструляцією. При цьому із бластодерми різними способами утворюються два шари клітин, або зародкові листки: зовнішній — ектодерма та внутрішній — ендодерма. Відомі чотири основні типи гаструляції (мал. 1.41): імміграція (переміщення), інвагінація (впинання), деламінація (розшарування) та епіболія (обростання). При імміграції ентодерма формується з клітин ектодерми переміщенням їх усередину бластули. Така гаструляція характерна для багатьох кишковопорожнинних. Інвагінація супроводжується впинанням вегетативного полюса всередину бластоцелю. Такий тип утворення гаструли характерний для членистоногих, ланцетників. Деламінація властива деяким медузам. При цьому два шари утворюються поділом клітин бластули паралельно її поверхні. Із внутрішніх клітин формується ендодерма, із зовнішніх — ектодерма. Епіболія виникає внаслідок більш інтенсивного поділу клітини анімального полюса та обростання ними клітин вегетативного полюса, які утворюють ентодерму, а зовнішні дрібні, що їх вкривають, — ектодерму. Такий тип гаструляції характерний для деяких червів. Описані типи гаструляції в чистому вигляді трапляються тільки в небагатьох тварин. У більшості тварин гаструляція змішана. Усередині гаструли утворюється порожнина — гастроцель (порожнина первинної кишки). Ця гастральна порожнина у більшості тварин згодом перетворюється на травний канал. Вона відкривається назовні бластопором (первинний рот). Тварини, в яких бластопор перетворюється на рот, називаються первинноротими (губки, кишковопорожнинні, черви, молюски, членистоногі). В інших тварин — вторинноротих рот формується з протилежного бластопору кінця, а останній перетворюється на анальний отвір. До вторинноротих належать голкошкірі й хордові.. Утворення гаструли Типи гаструляції (зверху — бластула, внизу — гаструла): а — інвагінація, б — епіболія, в — імміграція, д — деламінація

На наступній стадії утворення трьох зародкових листків — між екто- й ентодермою закладається третій зародковий листок — мезодерма. У більшості безхребетних (черви, молюски, членистоногі) вже під час дроблення утворюються дві-три клітини великих розмірів (телобласти, звідси телобластичний тип закладання). В процесі гаструляції вони розміщуються симетрично по боках первинної кишки на межі між екто- й ентодермою. Розмножуючись, вони дають клітини, що утворюють мезодермальні тяжі між екто- й ентодермою. Мезодерма в голкошкірих і хордових закладається за рахунок випинань двох кишенькоподібних порожнистих виростів ентодерми. Вони відшнуровуються і дають початок мезодермі. Зародкові листки хордових тварин внаслідок подальшого розвитку дають початок ембріональним зачаткам, з яких формуються тканини й органи: Стадія утворення трьох зародкових листків Формування органів із зародкових листків

Типи утворення мезодерми

В кожному зародковому листку продовжується подальше диференціювання клітин і виникають зачатки органів і тканин. Цей етап розвитку зародка називається органо- і гістогенезом. У всіх тварин певні органи беруть початок від одного й того самого зародкового листка. Одночасно з формуванням зачатків органів відбуваються спеціалізація клітин і утворення тканин — гістогенез. У рослин усі типи тканин мають єдине походження — з твірної тканини — меристеми. У тварин процеси гістогенезу складніші: тканини різних типів розвиваються з похідних різних зародкових листків. Важливу роль у цьому відіграють міжклітинні взаємодії, вплив біологічно активних речовин та неодночасна активність генів клітин. Першопричиною активації різних генів в ядрах клітин зародка на перших етапах його розвитку вважають різноякісність ділянок цитоплазми яйця. В зиготі гени репресовані. Спочатку відбувається активація генів, які контролюють утворення стовбурових (родоначальних) клітин, причому кожна із них в результаті розмноження утворює клон (ряд) клітин. Із певного числа клонів клітин потім виникають зачатки органів. Під час гісто- й органогенезу особливе значення мають міжклітинні впливи, які індукують розвиток різних тканин і органів. Речовини, що утворюються в одних клітинних системах, проникають у клітини інших систем і активують певні тканиноспецифічні гени, що зумовлює синтез ткани-носпецифічних білків і диференціації тканин. На пізніших етапах розвитку депресія генів відбувається переважно під впливом гормонів. У початково однорідній ектодермі ранньої гаструли під індукуючим впливом даху первинної кишки від покривної ектодерми відокремлюється нервова пластинка. По ходу нейруляції вона прогинається, утворюючи нервову трубку, яка відокремлюється від покривної ектодерми. Покривна ектодерма змикається над занурюваною нервовою трубкою . Розчленування мезодерми починається після відокремлення зачатка хорди, розміщеного по боках матеріалу сомітів — первинних сегментів, на які поділяються в черепно-хвостовому напрямку мезодермальні тяжі, та бічних несегментованих пластинок спланхнотома. Всередину соміти відокремлюють склеротомії, з яких потім утворюється хребет. З основної маси сомітів формуються міо-томи, які диференціюються в опорну мускулатуру. Окремі клітини майбутньої м'язової тканини (міобласти) зливаються на ранній стадії гістогенезу в багатоядерні синцитії, які як м'язові волокна з їхніми міофібрилами здійснюють на пізніших етапах розвитку скоротливі функції. На периферії в бік епідермісу соміти відділяють дерматоми, які є джерелом сполучнотканинної частини шкіри (дерми). Спланхнотом дає початок елементам видільної і статевої систем, утворює плевру, на якій закріплена кишкова трубка. З ентодерми розвивається травний канал. Вирости передньої ділянки кишки диференціюються в щитоподібну залозу та легеневий епітелій, а її середньої ділянки — в печінку й підшлункову залозу. Органогенез (формування органів) завершується здебільшого наприкінці ембріонального періоду розвитку. Проте диференціювання й ускладнення органів тривають і в постембріональний період. Тимчасові органи зародків і личинок тварин, які зникають у процесі їх розвитку, називаються провізорними. Вони забезпечують функції організму, який розвивається до формування й початку функціонування дефінітивних органів, властивих дорослим особинам. Такими органами є: черевні кінцівки і зябра личинок комах, зябра і хвіст пуголовків тощо. Диференціація тканин і органів під час зародкового розвитку (процес гістогенезу та органогенезу)

Закладання нервової трубкн: а: 1 — мезодермальний дах первинної кишки з бічними пластинками (2) розміщений між майбутньою нервовою пластинкою (3) та кишковою ентодермою (4), яка вкриває ще не замкнений просвіт кишок. З часом вона, зростаючи (стрілки вгору), закриває його; 5 — ектодерма; б — хорда по середній лінії даху первинної кишки відокремилася від решти мезодерми; в бічній мезодермі виникає целомічна щілина (б); ектодерма нервової пластинки (3) потовщена порівняно з ектодермою (5); в — краї нервової пластинки припідняті, утворюють нервові валики (7); 8 — хорда (просвіт кишок під нею замкнений); г — нервові гребені (9) зближуються й утворюють нервову трубку; д — нервова трубка (10) замкнена, відокремлена від ектодерми (5) та занурена під неї; первинні соміти (11) відокремлені від мезодерми, сформувалися стінки целома (12) із зовнішньою (соматоплевра — 73) і внутрішньою (спланхноплевра — 14) вистілками
")
Формування тканин та органів зародка (на прикладі ланцентника)

Сперматозоїд рухається по слизовій оболонці фалопієвих труб у пошуку яйцеклітини Проникнення сперматозоїду до яцеклітини крізь її оболонку

Повздовжній розріз крізь головку сперматозоїда. Генетичний матеріал міститься у голівці +8 днів. Вік ембріону — одна доба. Він розташувався в оптимальному місці на слизовій оболонці

+22 дні. Розвиток ембріону продовжуєетсья, і можна побачити утворення нервової трубки +24 дні. Один із найголовніших огранів організму — середце розвивається і починає битися

+9 тижнів. Судини на голові розвинулись у вигляді літери V ; починається розвиток кісток черепу + 10 тижнів. Розміром майже 3 см ембріон входить у зародковий період

+ 20 тижнів. Зародок вже розміром біля 20 сантиметрів. + 36 тижнів.

Насіння квіткових рослин

За нормальних умов розвитку окремі частини зародка й організму в цілому формуються узгоджено за місцем і часом. Це пояснюється тим, що зачатки одних органів розвиваються зазвичай під впливом взаємодій із зачатками, закладеними раніше. Явище взаємодії між частинами зародка під час ембріогенезу, за якого одна з них визначає напрямок розвитку сусідньої, дістало назву ембріональної індукції. Частина зародка, яка впливає на іншу, називається індуктором, або організатором. Згодом вона сама може виступати в цій ролі стосовно інших частин, які розвиваються пізніше. Питання про те, як відбувається процес індукції і якою є природа індуктора, поки що залишається нез'ясованим. Вважають, що індуктором є речовини білкової природи. Явище взаємодії частин зародка, що розвивається, та його біологічне значення Взаємовплив частин зародка (дія дорзальної губи бластопора як організатора): а — трансплантація; б — індуковане диференціювання другої нервової пластинки (вигляд зародка збоку); в — диференціювання трансплантата у тканини хорди, соміти тощо (поперечний переріз зародка); г — первинний і вторинний зародки; Бп — бластопор, 1НП — первинна і 2НП — вторинна нервові пластинки
 період може бути прямим і непрямим. За прям...")
Постембріональний (постнатальний) період може бути прямим і непрямим. За прямого розвитку після вилуплення із яйця або народження організм схожий на дорослу форму, тобто має основні морфологічні ознаки дорослої особини, але відрізняється від неї розмірами, пропорціями тіла й недорозвиненням деяких органів. Розвиток такого організму не супроводжується істотними перебудовами тіла. Цей тип розвитку властивий рибам, плазунам, птахам, ссавцям і багатьом безхребетним тваринам. Личинкова стадія відсутня. У прялеоліу.постембріональному розвитку розрізняють такі стадії: юність, зрілість і старість. Юність охоплює ювенільний період — від народження або вилуплення з яйця до початку статевого дозрівання. Період набуття статевої зрілості називається пубертатним. Старість — період затухання статевих функцій організму, який закінчується природною смертю. За непрямого постембріонального розвитку із зародкових оболонок виходить личинка, яка морфологічно відрізняється від дорослої форми, причому її розвиток супроводжується істотними перебудовами всього організму. Такий тип постембріонального розвитку називається розвитком з перетворенням, або метаморфозом. У природі він виявляється досить часто, причому у тварин, що перебувають на різних рівнях загальної організації, — у губок, кишковопорожнинних, голкошкірих, оболонників, круглоротих, деяких риб (вугри), земноводних. В одних випадках метаморфоз призводить до ускладнення організації (гідроїдні поліпи), в інших — до її спрощення (асцидії). В одних тварин відбувається докорінна перебудова організму (голкошкірі), а в інших будова організму принципово не змінюється (комахи). Розвиток може бути з повним і неповним метаморфозом. Післяембріональний (постембріональний) розвиток, його типи і етапи у тварин

У розвитку з повним перетворенням личинка різко відрізняється від дорослої тварини. Вона має личинкові (провізорні) органи, не властиві дорослій особині. Ріст і подальший розвиток личинки супроводжуються перебудовою всієї організації тіла. Розвиток з повним перетворенням відбувається у представників ряду Комахи (твердокрилі, лускокрилі, двокрилі, перетинчастокрилі, блохи), а також у,земноводних. У метаморфозі комах є три стадії: личинки, лялечки та імаго (доросла форма). Личинка в міру росту кілька разів линяє. Однак для перетворення на дорослу форму (імаго) вона має пройти стадію лялечки. У деяких комах лялечки рухливі, в інших — нерухомі. В цій стадії личинкові тканини і органи повністю перебудовуються. Більша частина органів руйнується. Зберігаються лише нервова система, зачатки статевих залоз та особливі утвори — іма-гінальні диски, за рахунок яких формуються органи дорослої форми, однак вони залишаються в сплюснутому, скрученому стані. Коли доросла форма залишає покриви лялечки, у сплюснуті імагінальні органи надходить кров, і вони випрямляються, набувають форми, характерної для дорослої комахи. Покриви комахи в цей час м'які й безбарвні, згодом у них з'являється хітин, вони твердіють і набувають характерного забарвлення. Так розвиваються лускокрилі (метелики), двокрилі, перетинчастокрилі та ін. Розвиток білана капустяного (1—S, повний метаморфоз) і сарани азіатської (6—13, неповний метаморфоз): 1 — самець; 2 — самка; 3 — кладка яєць; 4 — гусінь; 5 — лялечка; 6 — доросла крилата форма (імаго); 7 — кубишка з яйцями; 8 — яйце сарани; 9—13 — німфи (сарана піша)

Розвиток з неповним перетворенням спостерігається у представників прямокрилих, тарганів, бабок тощо. За неповного перетворення в комах, що вийшли з яйця, личинка має загальні риси будови дорослої форми (імаго), але відрізняється від неї розмірами, відсутністю крил, недорозвиненим статевим апаратом тощо). У личинок деяких комах є провізорні органи. Личинка кілька разів линяє (стадії німф) і без стадії лялечки після останньої линьки перетворюється на дорослу форму. У неї утворюються крила, дозрівають статеві залози, з'являються зовнішні статеві придатки. Розвиток з неповним перетворенням спостерігається також у кліщів. Розвиток із перетворенням має ряд переваг перед прямим способом розвитку. Личинка, як правило, використовує джерело їжі, відмінне від дорослої особини, що приводить до послаблення внутрішньовидової конкуренції за ресурси. У губок, кишковопорожнинних, багатощитинових червів личинка рухома і слугує для розселення виду. Личинки деяких видів (ракоподібних, павукоподібних, земноводних) здатні розмножуватись. Ця властивість дістала назву неотенії. Здатність до неотенії має пристосувальне значення для тих видів тварин, у процесі індивідуального розвитку яких відбувається зміна середовища проживання. Якщо умови існування дорослої стадії украй несприятливі, збільшення кількості личинок підвищує шанси популяції на виживання. Старіння і смерть організмів Старіння — закономірний процес вікових змін, що веде до зниження адаптаційних можливостей організму, збільшення ймрвірності смерті. Старіння властиве всім організмам і перебігає на молекулярно-генетичному, клітинному, тканинному, органному рівнях організації живого.. Смерть — припинення життєдіяльності організму. Основним біологічним сенсом смерті можна вважати підтримання колообігу речовин та енергії в біосфері.
Схожі презентації











Категорії


")

")






