Презентація на тему:
ВУГЛЕВОДИ: Класифікація; хімічні і фізичні властивості; структура моно-, оліго- та полісахаридів.
Завантажити презентацію





























-O- -D-Глюкоріранозилуронова кислота(1,3)-2-ацетиламідо-2-дезокси- -D-гл...")

-O- -D-глюкопіранозилуронова кислота-2-сульфо-(1,4)-2-сульфоамі...")
























 вперше виявлені в 1968 р. швейц...")




, тимін (Т) і цитозин (С): урацил (U) 2...")







 Рідинний хроматограф фірми")

 та дезоксирибонуклеїнові (ДНК) кислоти Нуклеїнові кислот...")



ВУГЛЕВОДИ: Класифікація; хімічні і фізичні властивості; структура моно-, оліго- та полісахаридів.
Завантажити презентаціюПрезентація по слайдам:

ТЕМА: ВУГЛЕВОДИ: Класифікація; хімічні і фізичні властивості; структура моно-, оліго- та полісахаридів. Лектор: асис. Руснак Н. І.

Вуглеводи. Біологічні функції вуглеводів. Класифікація вуглеводів. Моносахариди: класифікація, будова молекули, властивості. Олігосахариди: будова, біологічні функції. Гомо- та гетерополісахариди: будова молекули, біологічні функції. План

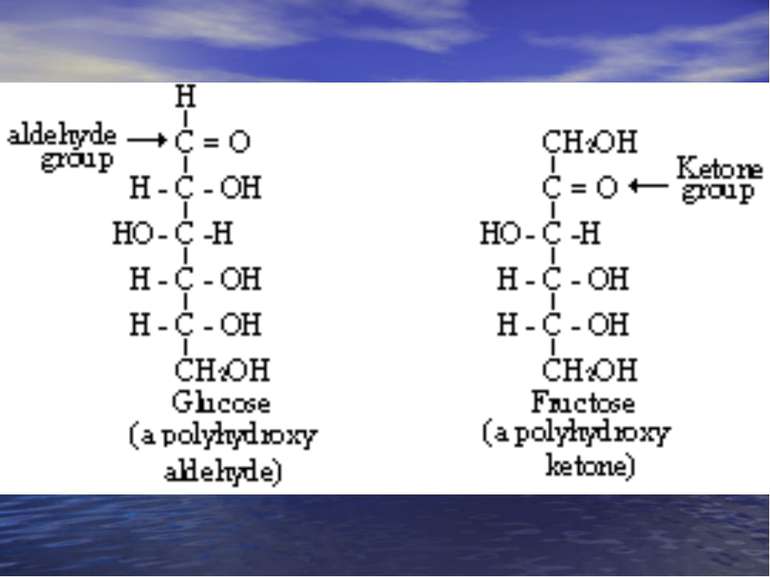
Багато простих вуглеводів мають емпіричну форму СnН2nОn. Було доказано, що вони складаються з Карбону, Оксигену і Гідрогену, при чому співвідношення між воднем і киснем таке як у воді, тому їх назвали “вуглеводи” – “гідрати карбону” Сn(Н2О)n Вуглеводи це численна група природних i синтетичних сполук, які є за хімічною будовою полігідроксильними речовинами, що містять альдегідну або кетонну групи, або ж утворюючими їх при гідролізі.

Біологічні функції вуглеводів Енергетична – при окисленні 1 грама вуглеводів виділяється 17,2 кДж/г Резервна: глікоген. Вуглеводи є джерелом Карбону з якого синтезуються інші речовини (білки, ліпіди, нуклеїнові кислоти). Вуглеводи є структурними одиницями ДНК і РНК (рибоза дезоксирибоза). Олігосахаридні комплекси глікопротеїни і гліколіпіди мембран утворюють центри розпізнавання біомолекул, виконують роль антигенів.

Класифікація вуглеводів Моносахариди – прості вуглеводи, полігідроксиальдегіди і полігідроксикетони. Наприклад: глюкоза, рибоза, ксилоза, фруктоза Олігосахариди – вуглеводи молекули яких містять 2-10 моносахаридних залишки. Наприклад сахароза, лактоза, маноза, рафіноза.. Полісахариди – вуглеводи молекули яких містять велику кількість моносахаридних залишків Наприклад: целюлоза, крохмаль, глікоген, гепарин.

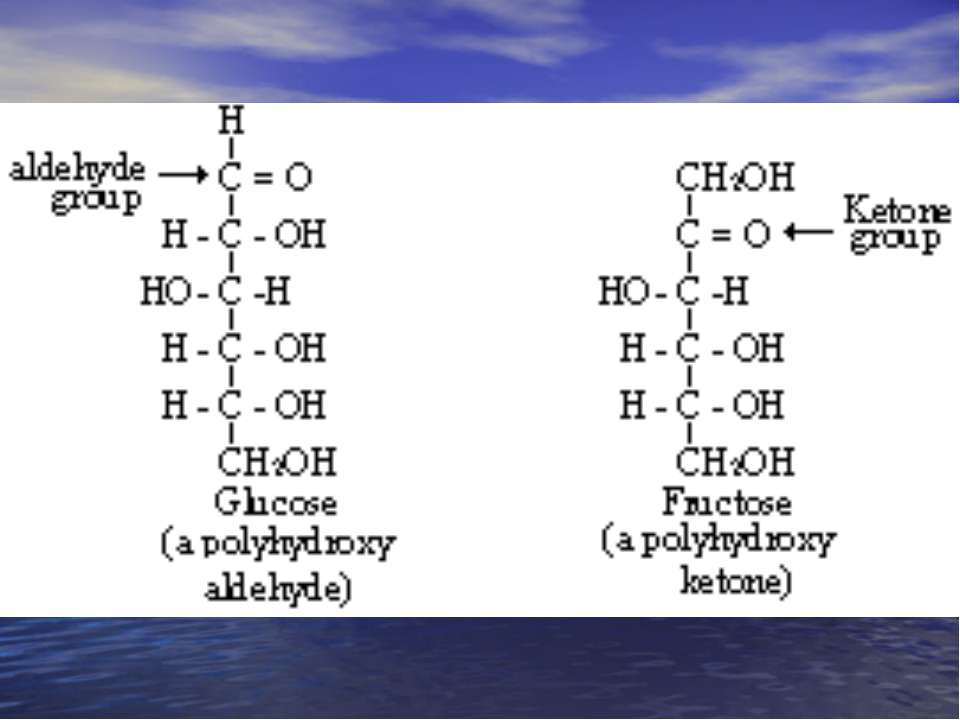
Класифікація моносахаридів. За природою карбонільної групи: Кетози – містять кетонну і декілька гідроксильних груп. Альдози – містять альдегідну і декілька гідроксильних груп. За кількістю атомів Карбону: Тетрози; Пентози; Гексози; Гептози.

Гліцериновий альдегід.

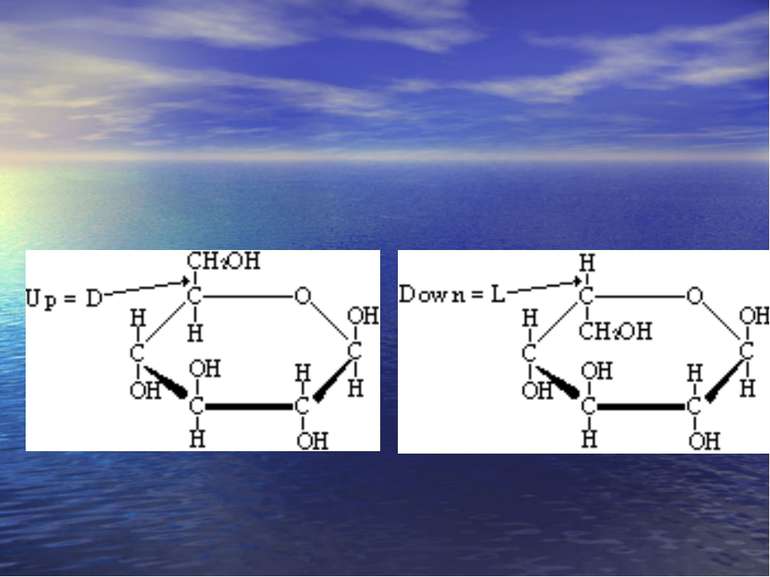
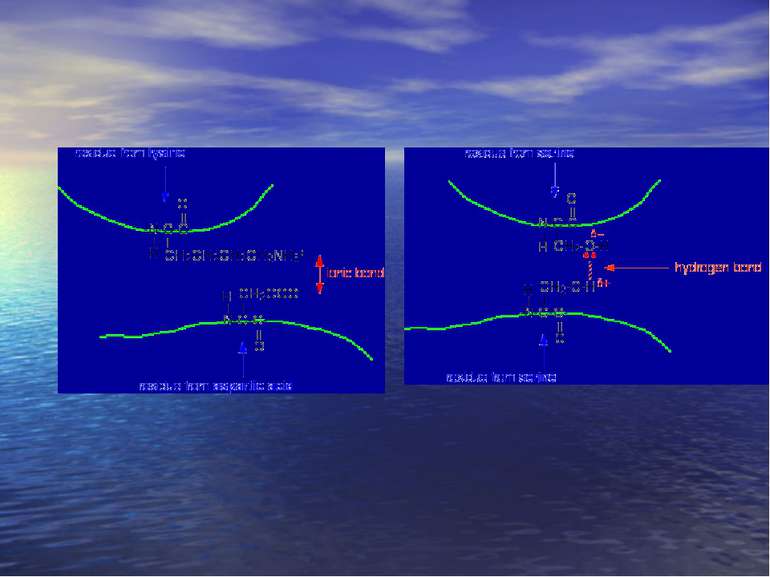
Стереоізомерія – явище існування декількох просторових ізомері: геометрична, оптична. Оптичні ізомери – це сполуки, які мають один або декілька асиметричних (хіральних) атоми карбону. Енантіомери: – Це просторові ізомери, які відносяться один до одного як предмет і його дзеркальне відображення (D і L конфігурація). – вони повертають площину поляризованого світла в різних напрямках: правоповертаючі – за годинниковою стрілкою (+) і лівоповертаючі (-) – проти годинникової стрілки енантіомери. Діастаріоізомери – оптичні ізомери, які не є дзеркальними відображеннями один одного. Епімери – це оптичні діастеріоізомери, які відрізняються положенням функціональних груп лише біля одного асиметричного атома.

Альдози

Кетози

Важливі моносахариди. D-Галактоза D-Глюкоза D-Фруктоза D-Рибоза 2-Дезокси-D-рибоза
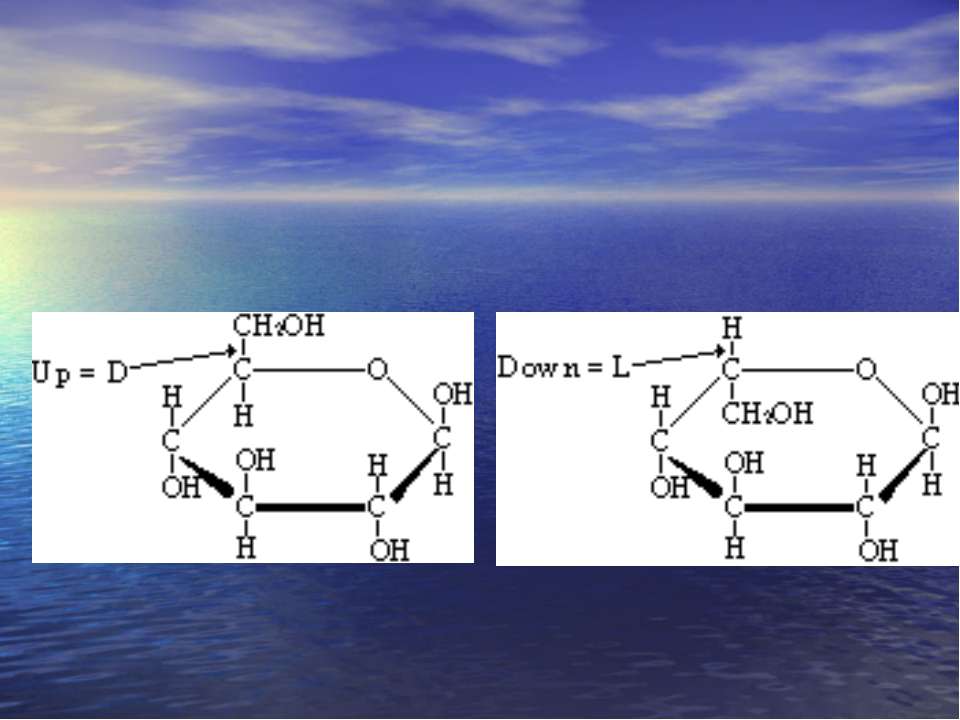

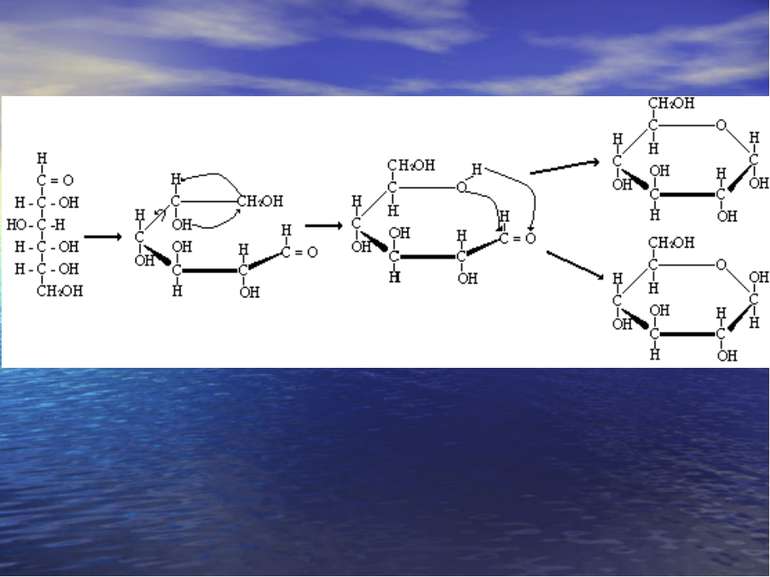
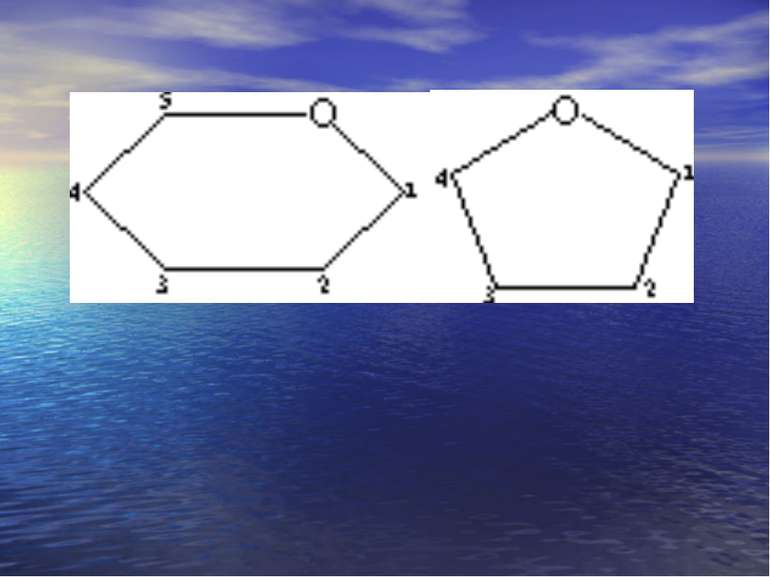
-D-Моносахариди -D- Моносахариди

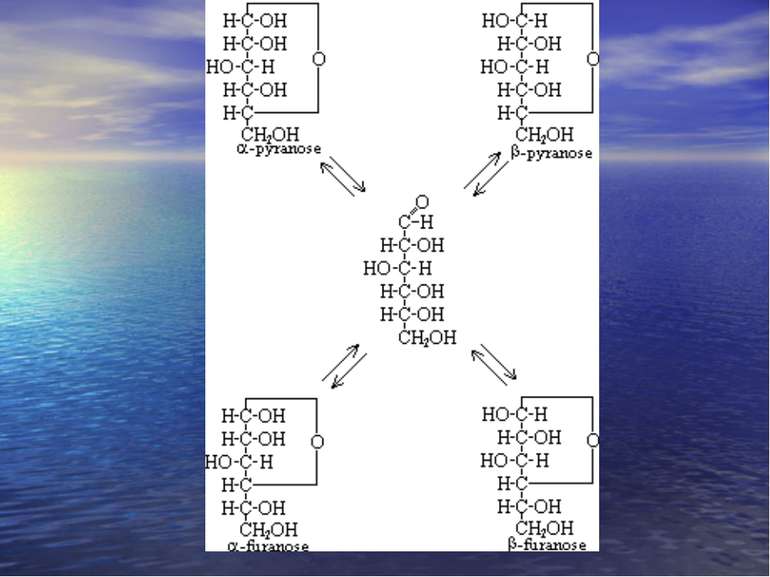
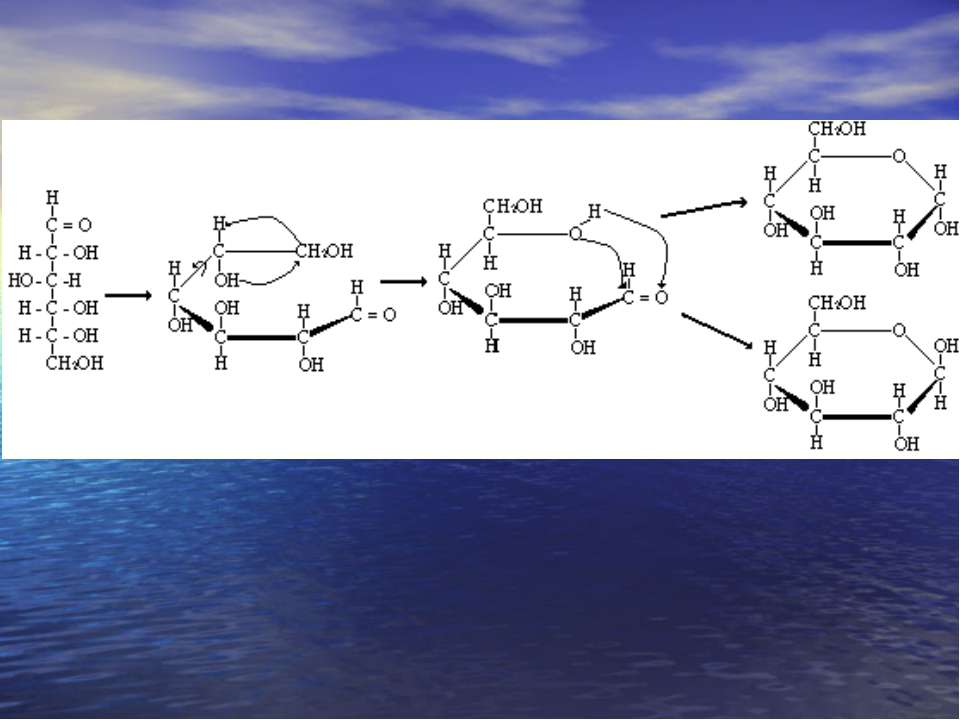
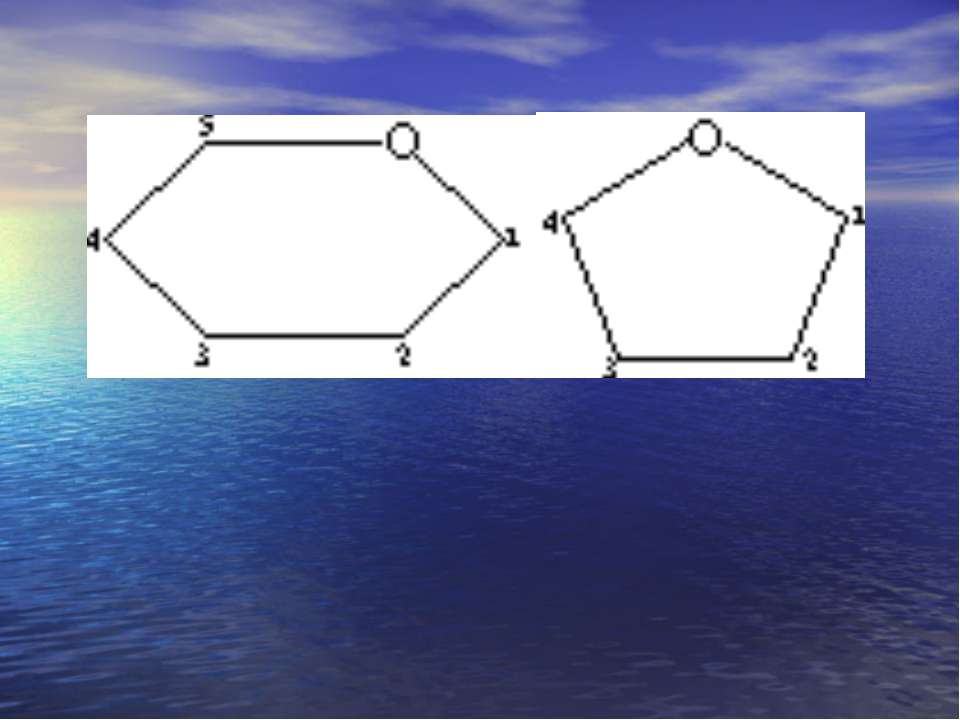
Хеуорса і Фішера проекції -форма -форма

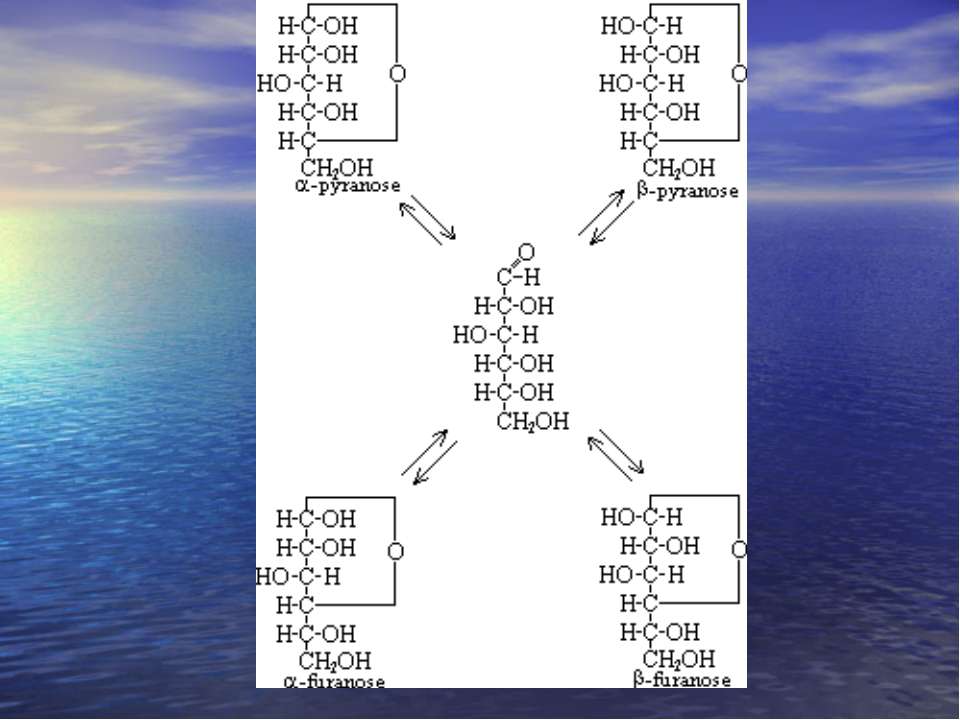
Мутаротація. При розчиненні моносахаридів у воді циклічні -, -фуранозні, піранозні форми і альдозна форми переходять одна в одну. Це явище цикло-оксо-таутомерією. Цей процес супроводжується спонтанною змінною величини кута повороту площини поляризованого світла називається мутаротацією,

Окислення При окисленні моносахаридів слабкими окисниками утворюються альдонові (гліконові) кислоти. При окисленні глюкози утворюється глюконова кислота.

Сильні окисники окисляють моносахариди до дикарбонових кислот (альдарових – цукрових кислот) глюкоза окисляється до гюкарової кислоти.

В біологічних процесах ферменти здатні окислити первину спиртову групу моносахаридів до карбоксильної при цьому утворюються уронові кислоти для глюкози глюкурону кислоту.

Відновлення. При відновленні альдоз і кетоз утворюються багатоатомні спирти (цукрові спирти). Глюкоза відновлюється до сорбітолу.

Глікозиди. Метил- -D-глюкозид Метил- -D- глюкозид

Естери -D-глюкозо-1-фосфат -D-Глюкозо-6-фосфат

Аміно цукри -D-Глюкозамін -D-Галактозамін N-Ацетил -D-глюкозамін

Ізомеризація

Олігосахариди - вуглеводи, під час гідролізу яких утворюється ввід 2 до 10 моносахариди. Дисахариди C12H22O11 – білі кристалічні речовини, солодкі на смак, добре розчині у воді, під час гідролізу яких утворюються два моносахариди. Відновлюючі і невідновлюючі.

Реакція утворення дисахаридів Моносахарид + Моносахарид = Дисахарид + Н2O (Як напівацеталь (Як спирт) (Глікозит ) напівкеталь)

Лактоза Мальтоза

Сахароза – не відновлюючий цукор

Інвертний цкор – еквівалентна кількість глюкози і фруктози, в розчині який утворюється при гідролізі сахарози

Амілопектин Целюлоза Амілоза Амілоза
-O- -D-Глюкоріранозилуронова кислота(1,3)-2-ацетиламідо-2-дезокси- -D-гл...")
(1,4)-O- -D-Глюкоріранозилуронова кислота(1,3)-2-ацетиламідо-2-дезокси- -D-глюкопіраноза Гіалуронова кислота - містить N-ацетилглюкозамін і глюкуронову кислоту зв'язані (1 - 3) і (1-4) глікозидними зв'язками . Є основою сполучної тканини, міститься у синовіальній рідині, шкірі, хрящах, скловидному тілі ока, пупковині. Синовіальна рідина містить 0.02 – 0.05% гіалуронової кислоти.

Хондроітин сульфат. Складається N-ацетил глюгозамін-6 сульфату і глюкуронової кислоти або L-ідуронової кислоти. (1,4)-O- -D-глюкопіранозилуронова кислота-(1,3)-2-ацетоаміно-2-дезокси-6-O-сульфо- -D-галактопіраноза
-O- -D-глюкопіранозилуронова кислота-2-сульфо-(1,4)-2-сульфоамі...")
Гепарин. (1,4)-O- -D-глюкопіранозилуронова кислота-2-сульфо-(1,4)-2-сульфоамідо-2-дезокси-6-O-сульфо- -D-глюкопіраноза

АМІНОКИСЛОТИ, ПЕПТИДИ, БІЛКИ.

У м'язах, легенях, селезінці, нирках на білки припадає більше 70-80 % сухої маси; в печінці – 57 %, у мозку – 45 %. Найнижчий вміст білка в кістці і у зубах – 20 і 18 %. Неоднаковий вміст білка і у різних субклітинних органелах. Найбільше білка в гіалоплазмі (внутрішньоклітинний сік). Якщо прийняти загальний білок клітини за 100 %, то на гіалоплазму припадає 40 %. Мітохондрії та мікросоми містять по 20 %, ядро – 12 %, лізосоми – 2 %, пероксисоми – 2,5 %, плазматична мембрана – 1,5 % білка.

АМІНОКИСЛЛТИ

За біологічним значенням Замінні; Незамінних; Абсолютно незамінних амінокислот є 8: валін, лейцин, ізолейцин, треонін, лізин, метіонін, фенілаланін і триптофан


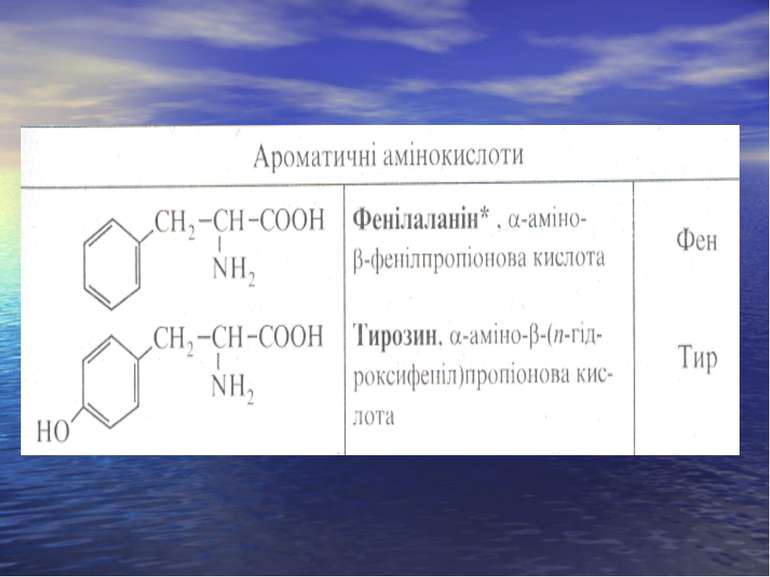
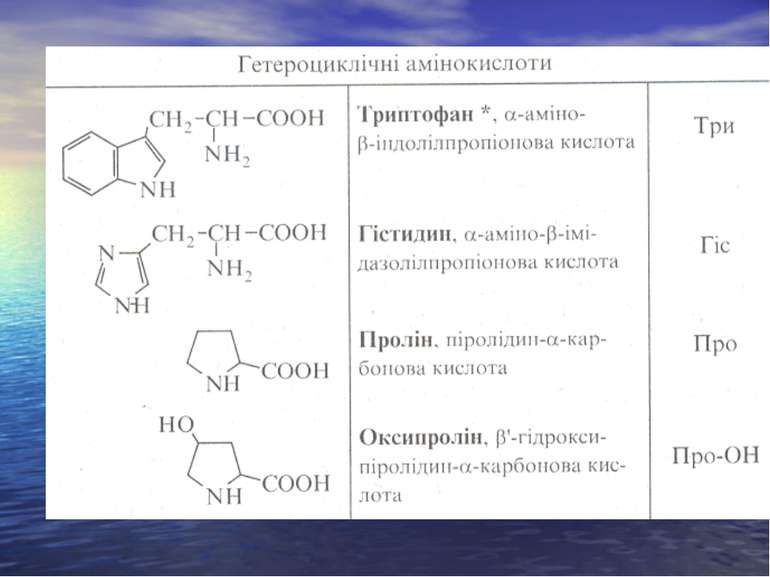
СІРКОВМІСНІ АМІНОКИСЛОТИ
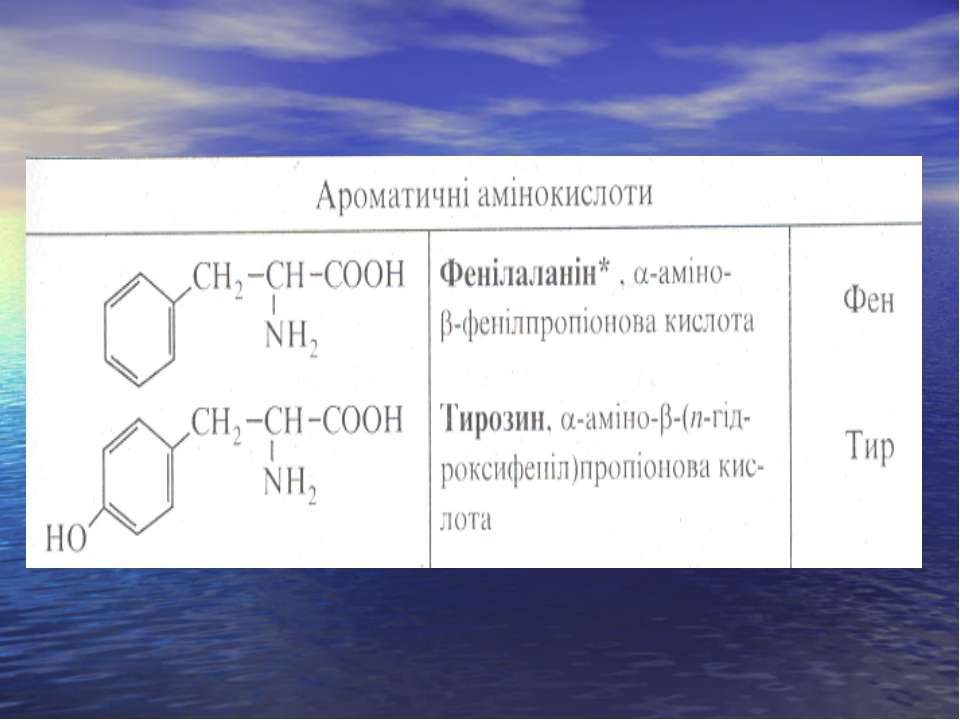
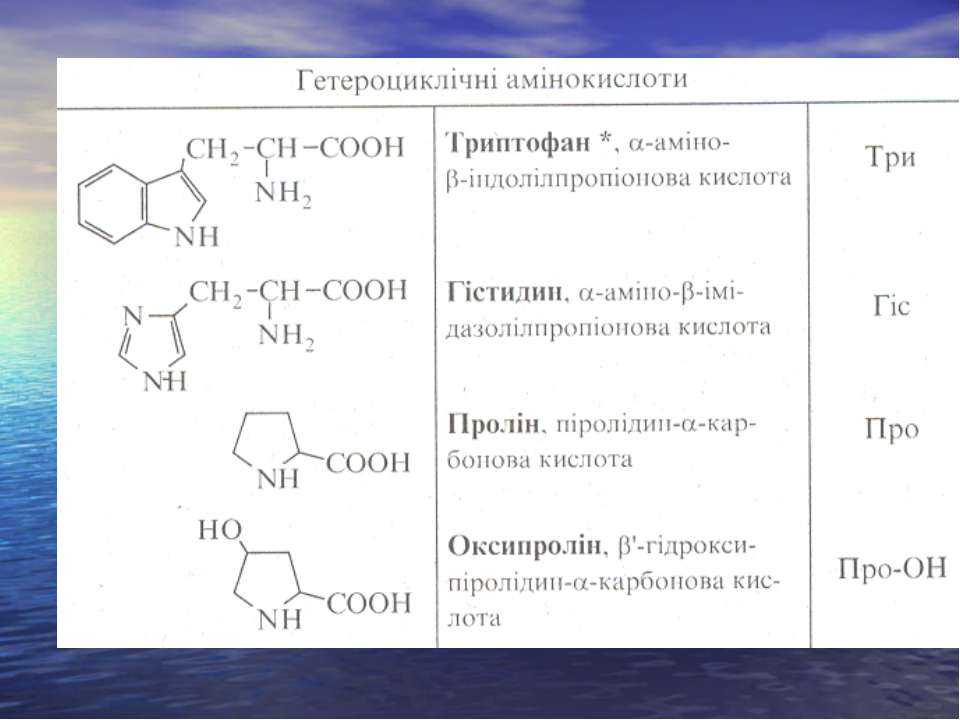

ОПТИЧНА КОНФІГУРАЦІЯ

КИСЛОТНІСТЬ

Утворення амідів: Утворення солей:

Декарбоксилювання

Утворення комплексних сполук

Утворення естерів

Взаємодія з карбонільними сполуками

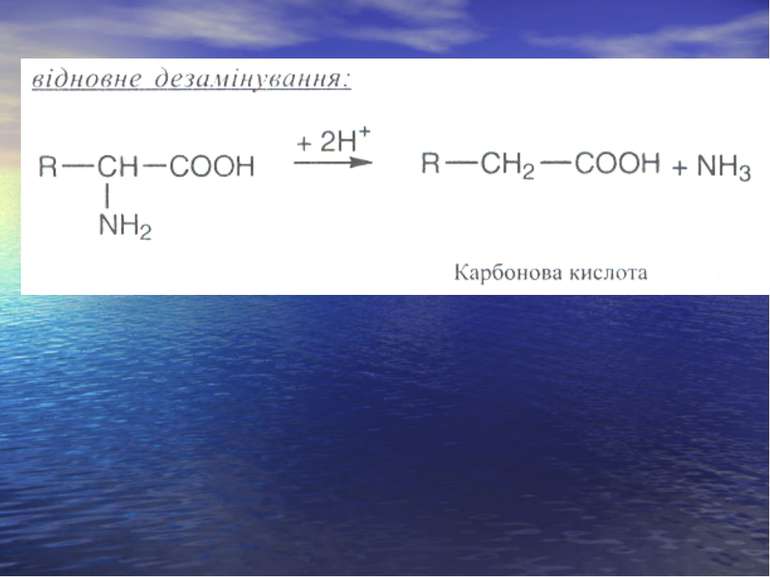
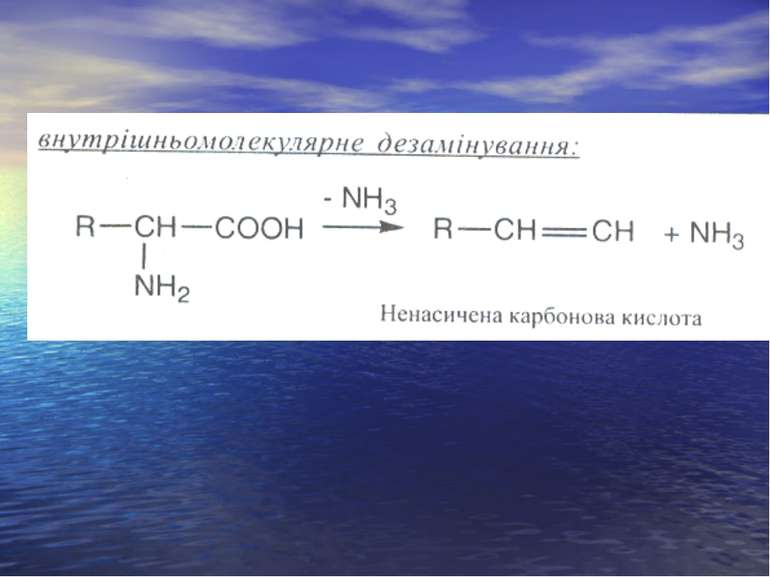
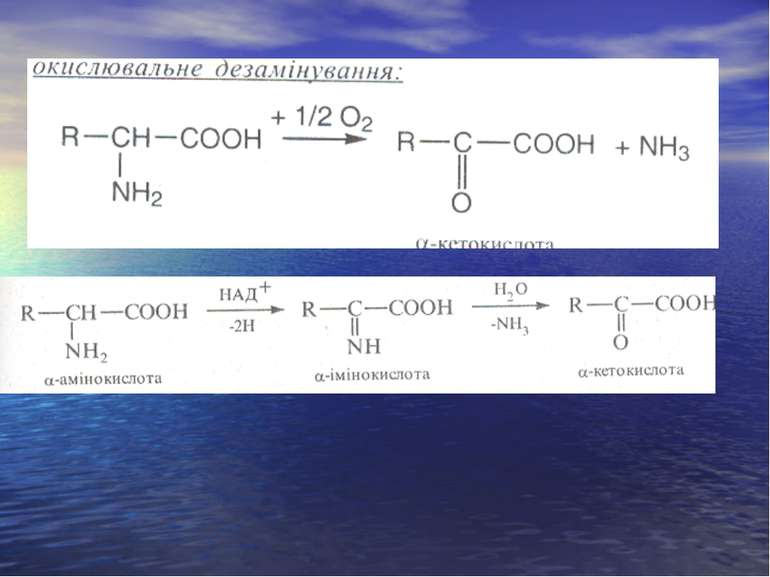
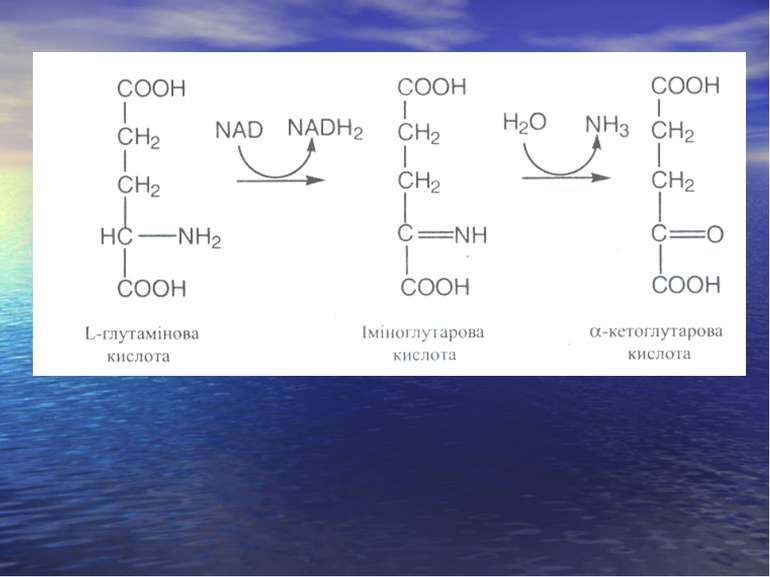
Дезамінування
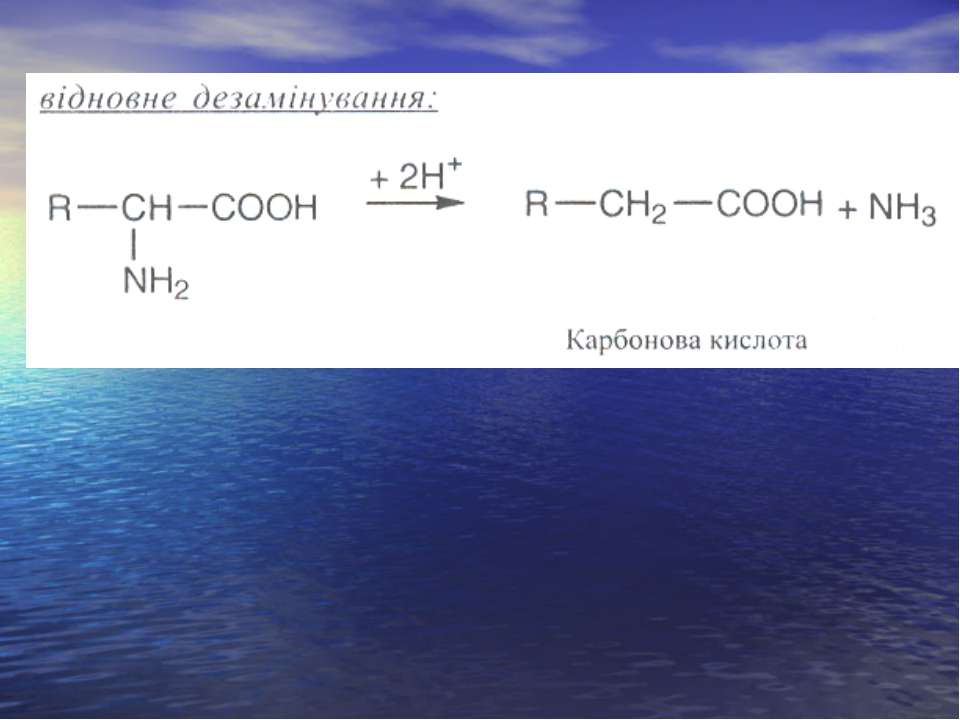
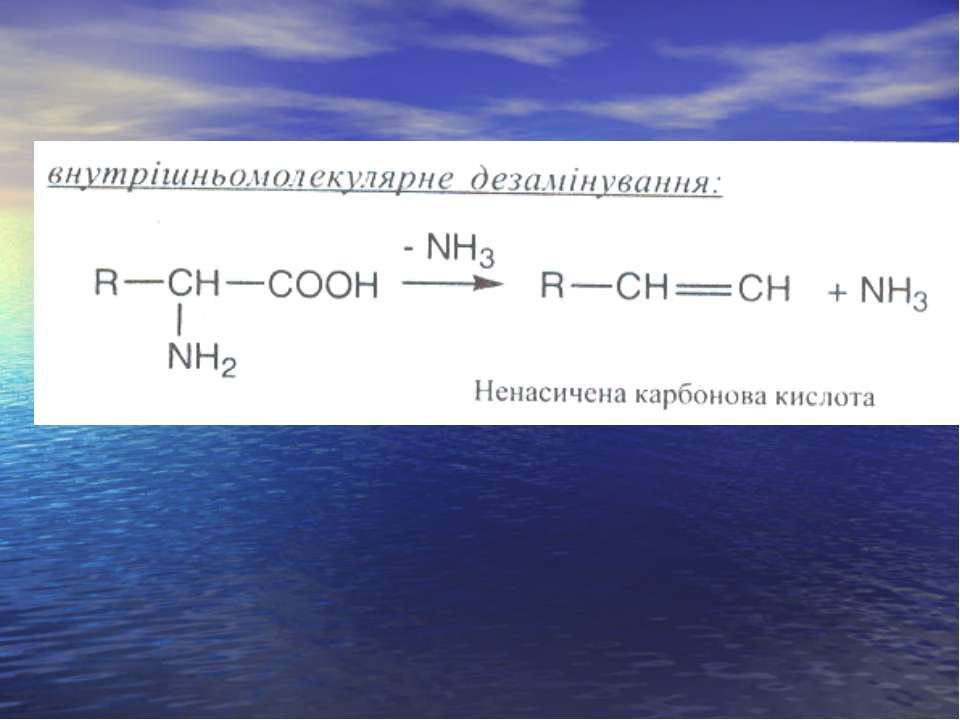
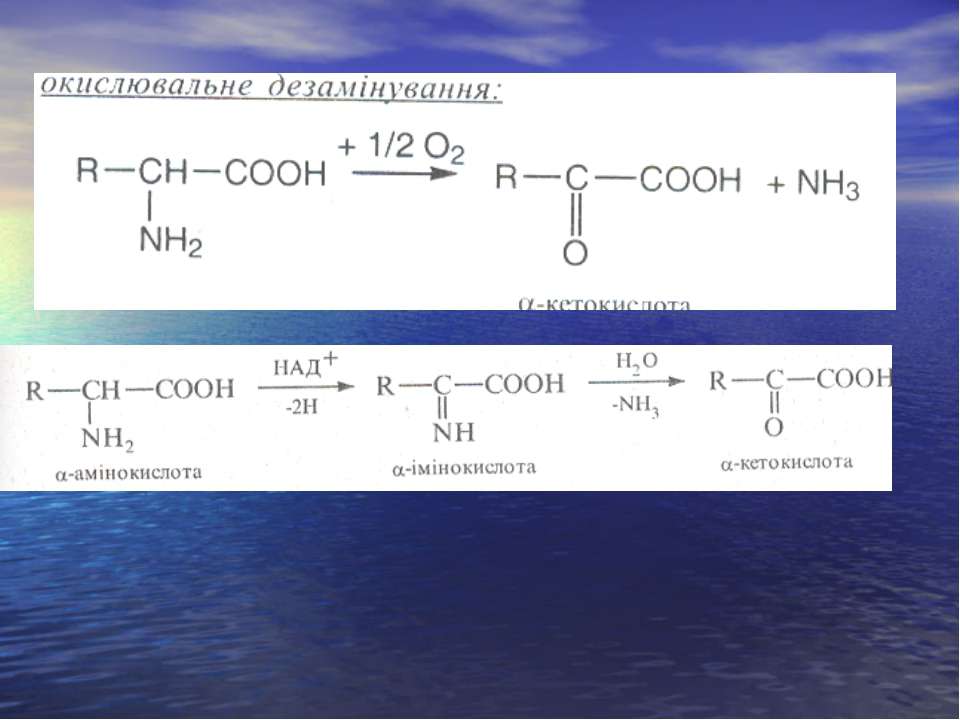
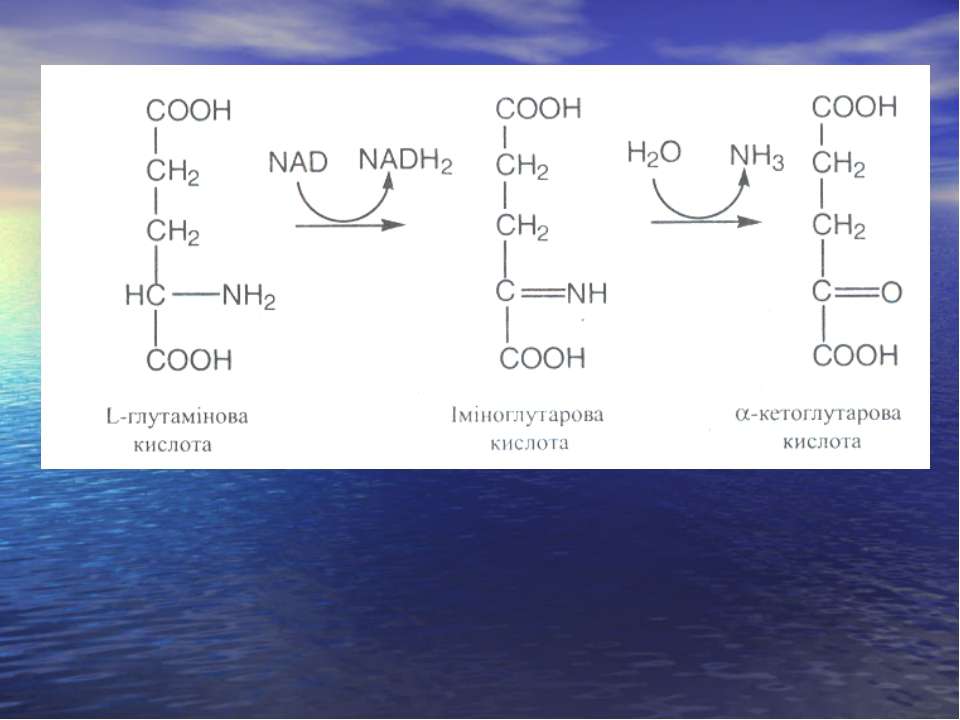

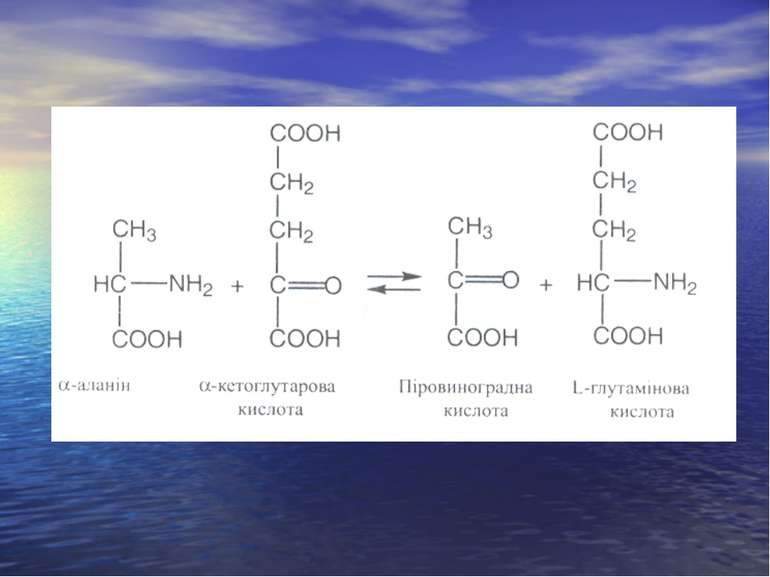
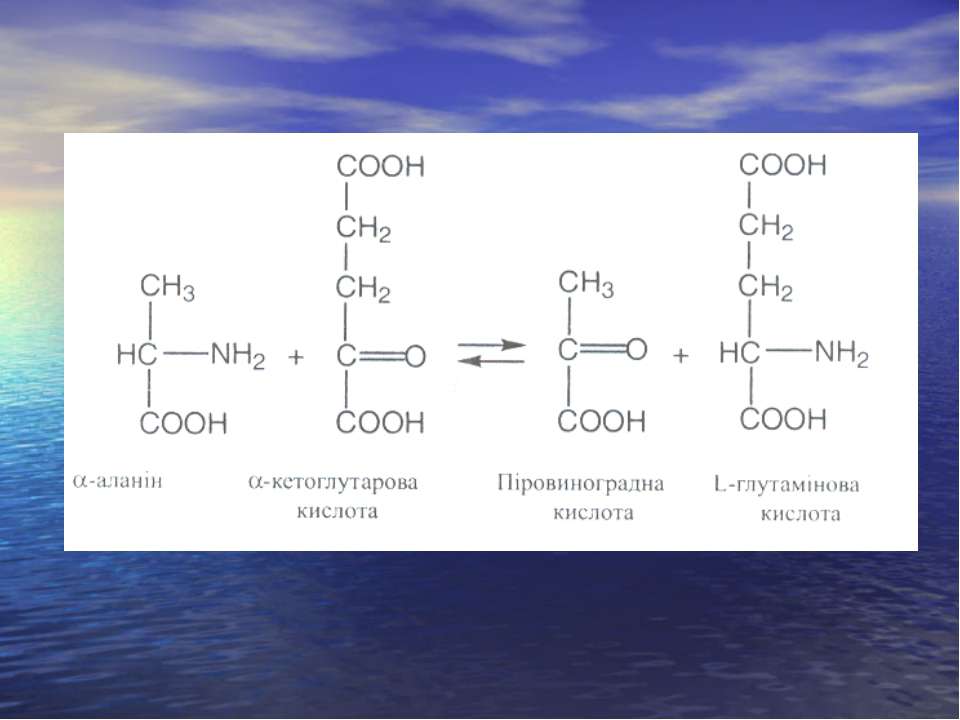
Реакція трансамінування

Утворення пептидів

Якісні реакції на амінокислоти

Первина структура

Вторинна структура

Альфа спіраль

Бета структура

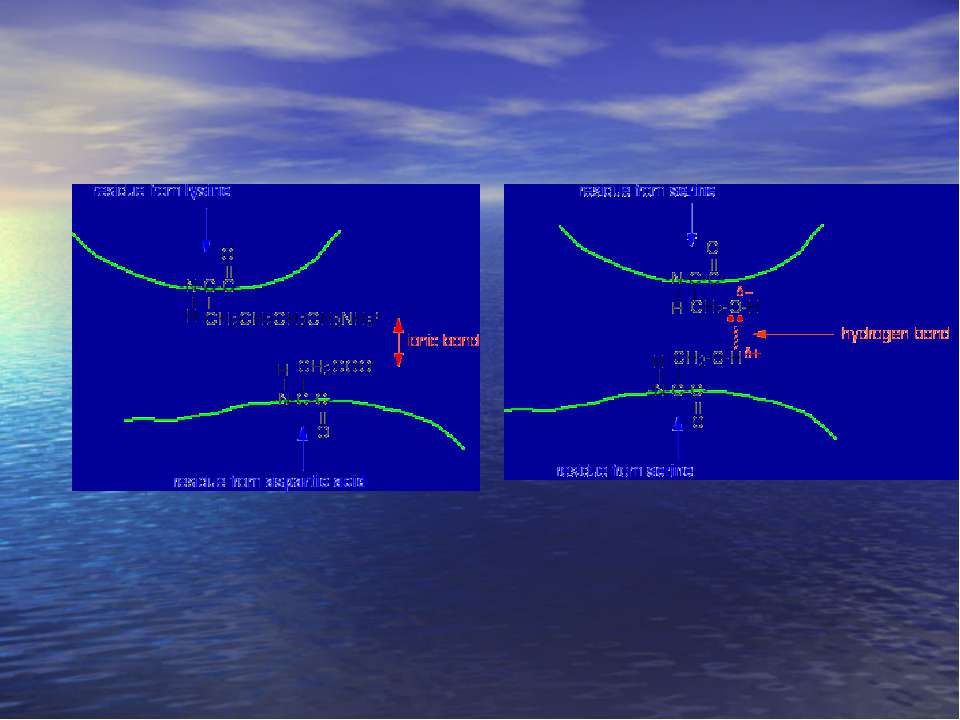
Зв'язки, що формують третину стркктуру білка

ГЕМОГЛОБІН

Нуклеїнові кислоти
 вперше виявлені в 1968 р. швейц...")
Нуклеїнові кислоти (від лат. nucleus— «ядро») вперше виявлені в 1968 р. швейцарським хіміком Ф. Мішером в ядрах клітин. Пізніше аналогічні речовини були знайдені також у протоплазмі клітин. Нуклеїнові кислоти забезпечують зберігання й передачу спадкової інформації, беручи безпосередню участь у синтезі клітинних білків. Вони входять до структури складних білків нуклеопротеїдів, які містяться в усіх клітинах організму людини, тварин, рослин, бактерій та вірусів. Кількість нуклеїнових кислот у різних нуклеопротеїдах, крім вірусних, коливається в межах від 40 до 65%

Будова нуклеїнових кислот Нуклеїнові кислоти, подібно до білків, являють собою високомо-лекулярні органічні сполуки, проте на відміну від білків, які утворю ють при гідролізі α-амінокислоти, мономерними одиницями нуклеїнових кислот є нуклеотиди. Тому нуклеїнові кислоти називають ще полінуклеотидами. Мономери нуклеїнових кислот — нуклеотиди — мають також доволі складну будову. При гідролізі нуклеотидів утворюються вуглевод, ортофосфорна кислота та гетероциклічні основи.

Таким чином, в цілому гідроліз нуклеїнових кислот можна подати у вигляді схеми:

Будова нуклеїнових кислот У залежності від природи вуглеводу, що входить до складу нуклеотидів, нуклеїнові кислоти розподіляють на два види дезоксирибонуклеїнові кислоти (ДНК), які містять вуглевод 2-дезокси-D-рибозу, та рибонуклеїнові кислоти (РНК), які вміщують вуглевод D-рибозу. 2-дезокси-D-рибоза та D-рибоза знаходяться в нуклеїнових кислотах у β-фуранозній формі: в 2-дезокси- D-рибоза D-рибоза

Гетероциклічні основи, що входять до складу нуклеїнових кислот, є похідними пурину та піримідину. До основ групи пурину відносяться аденін (А)* і гуанін (G): Аденін (А) 6-амінопурин Гуанін (G) 2-аміно-6-оксопурин
, тимін (Т) і цитозин (С): урацил (U) 2...")
Основами групи піримідану , урацил (U), тимін (Т) і цитозин (С): урацил (U) 2,4- діоксипіримідин Тимін (Т) 5-метил-2,4- діоксипіримідин Цитозин (С) 4-аміно-2-оксипіримідин

До складу ДНК входять аденін, гуанін, цитозин і тимін. а до скла ду РНК аденін, гуанін, цитозин і урацил. Для гуаніну, урацилу, тиміну та цитозину рпягтива лактам-лактимна таутомерія:

У нуклеїнових кислотах органічні основи сполучені N-глікозидним зв'язком з залишком D-рибози або 2-дезокси-D-рибози. Глікозидний зв'язок здійснюється за участю напівацетального гідроксилу моносахариду (С1') . N-Глікозиди, що складаються з залишків нуклеїнових основ і D-рибози або 2-дезокси-D-рибози, називають нуклеозидами У залежності від природи вуглеводного залишку розрізняють рибонуклеозиди та дезоксирибонуклеозиди. N-глікозидний звязок

аденін дезоксирибоза дезоксирибонуклеозид N-глікозидний звязок

Назви нуклеозидів утворюють аналогічно назвам глікозидів. Так, нуклеозид, який складається з рибози та урацилу, називають β-урацилрибофуранозидом, нуклеозид з дезоксирибози і аденіну β-аденіндезоксирибофуранозидом тощо. Проте, частіше застосовують назви, котрі для рибонуклеозидів утворюють з тривіальних назв відповідних нуклеїнових основ із закінченням -идин (-ідин) у піримідинових і -озин у пуринових нуклеозидів, наприклад аденозин, гуанозин, цитидин і уридин:

У назвах дезоксирибонуклеозидів додатково вводиться префікс дезокси- (дезоксі-), наприклад: дезоксіаденозин, дезоксигуанозин, дезоксицитидин. Винятком є назва нуклеозиду, що складається з дезоксирибози та тиміну тимідин (замість дезокситимідину). тимідин дезоксіаденозин дезоксіцитидин дезоксигуанозин

У нуклеїнових кислотах гідроксильна група біля С5' або С3' пентозного залишку нуклеозиду етерифікована ортофосфорною кислотою. Складний ефір фосфорної кислоти (фосфат) нуклеозиду назива ють нуклеотидом. У залежності від природи пентози розрізняють рибонуклеотиди та дезоксирибонуклеотиди.

Номенклатура нуклеотидів Назви нуклеотидів як монофосфатів Назви нуклеотидів як кислот Аденозин-5'-монофосфат (АМФ) 5'-аденілова кислота Гуанозин-5'-монофосфат (ГМФ) 5'-гуанілова кислота Цитидин-5'-монофосфат (ЦМФ) 5'-цитидилова кислота Уридин-5'-монофосфат (УМФ) 5'-уридилова кислота Дезоксіаденозин-5'-монофосфат (ДАМФ) дезоксіадепілова кислота Дезоксигуанозин-5'-моІюфосфат(ДГМФ) дезоксигуанілова кислота Дезоксицитидип-5'-моІюфосфат (ДЦМФ) дезоксицитидилова кислота Тимідин-5'-монофосфат (ДТМФ) тимідилова кислота
 Рідинний хроматограф фірми")
2. Високоефективна рідинна хроматографія (ВЕРХ) Рідинний хроматограф фірми

Нуклеїнові кислоти являють собою продукти полімеризації моно-нуклеотидів. Нуклеотиди сполучаються в довгі ланцюги за допомогою фосфодіефірних зв'язків, які утворюються за участю гідроксилу при С*' попередньої нуклеотидної ланки та гідроксилу, що належить С ' дальшої нуклеотидної ланки. Мононуклеотиди, їх похідні та динуклеотиди присутні в клітинах також у вільному вигляді та виконують важливу роль в обміні речо-вин. В усіх тканинах організму, поряд з нуклеозидмонофосфатами, містяться ди- та трифосфати нуклеозидів. Особливо широко відомі аденозин-5’- фосфат (АМФ), аденозин-5’-дифосфат (АДФ) і аденозин-5'-трифосфат (АТФ). Ці нуклеотиди здатні до взаємоперетворення шляхом фосфорилю-вання (приєднання одного або двох залишків фосфорної кислоти до АМФ) або ж дефосфорилювання (відщеплення одного або двох за лишків фосфорної кислоти від АТФ). При дефосфорилюванні виділя ється значна кількість енергії, котра використовується в організмі для проходження тих чи інших біологічних процесів, наприклад, у біо синтезі білка.
 та дезоксирибонуклеїнові (ДНК) кислоти Нуклеїнові кислот...")
Рибонуклеїнові (РНК) та дезоксирибонуклеїнові (ДНК) кислоти Нуклеїнові кислоти являють собою високомолекулярні гетерополімери, які складаються з залишків ортофосфорної кислоти та рибози або дезоксирибози, що чергуються, сполучених з нуклеїновими основами, котрі виступають у полімерному лан цюзі як «бокові групи» Схема ділянки полінуклеотидного ланцюга молекули нуклеїнової кислоти

Певна послідовність нуклеотидних ланок у полінуклеотидному ланцюзі називається первинною структурою нуклеїнових кислот. (Просторова орієнтація полінуклеотидних ланцюгів у молекулі називається вторинною структурою нуклеїнових кислот. Вперше вторинну структуру ДНК у вигляді моделі з подвійної спіралі описали американський біохімік Дж. Уотсон і англійський біохімік Ф. Крик (1953 p.). Узагальнивши роботи Л. Полінга, А. Тод-да, Е. Чаргаффа, М. Уілкінса та інших, вони дійшли висновку, що молекула ДНК являє собою дві паралельні правозакручені спіралі (подвійна спіраль), фіксовані між собою ван-дер-ваальсовими силами притягання, що діють вздовж спіралі між ядрами нуклеїнових основ (міжплощинна вертикальна взаємодія). Крім того, вторинна структу ра стабілізується водневими зв'язками між залишками нуклеїнових основ двох паралельних спіралей.

За моделлю Уотсона та Крика діаметр спіралі 1,8-2,0 нм. Кожний виток спіралі міс тить 10 пар основ. Крок спіралі складає 3,4 нм. Відстань між площинами основ по вертика лі дорівнює 0,34 нм. Полінуклеотидні ланцюги нм подвійної спіралі розташовані в протилежних напрямках. На одній нитці подвійної спіралі фосфодіефірні зв'язки утворені за типом 5'-3', а на другій навпаки, за типом 3'-5'.Між піримідиновими та пуриновими нук леїновими основами паралельних ниток под війної спіралі ДНК утворюються водневі зв'язки. При цьому аденін утворює зв'язок з тиміном, а гуанін з цитозином. Тому їх називають комплементарними парами (AT і GC). РНК являє собою одинарну спіраль. Вторинна структура РНК має відносно невелику масу. Відомі три типи РНК: матрична РНК (мРНК), або інформаційна РНК, рибосомальна РНК (рРНК) і транспортна РНК (тРНК). Схема подвійної спіралі ДНК

Нитка А Нитка В Комплементарні пари нуклеїнових основ у складі подвійної спіралі ДНК
Схожі презентації











Категорії











