Презентація на тему:
Міжклітинна комунікація
Завантажити презентацію


 ек...")

















Міжклітинна комунікація
Завантажити презентаціюПрезентація по слайдам:

Міжклітинна комунікація
 ек...")
Взаємодія між клітинами в організмі відбувається за трьома механізмами: а) екстраклітинна взаємодія - опосередковано гормонами, ростовими факторами, нейротрансміттери та цитокіни, б) внутріклітинна взаємодія через іншу групу посередників (Са, рН, кераміди, оксид азоту,с-АМР, реактивний кисень), активує системи сигнальної трансдукції і модулює в)міжклітинну взаємодію через gap junction channels (3), яка відбувається через субклас міжклітинних комунікаційних молекул.
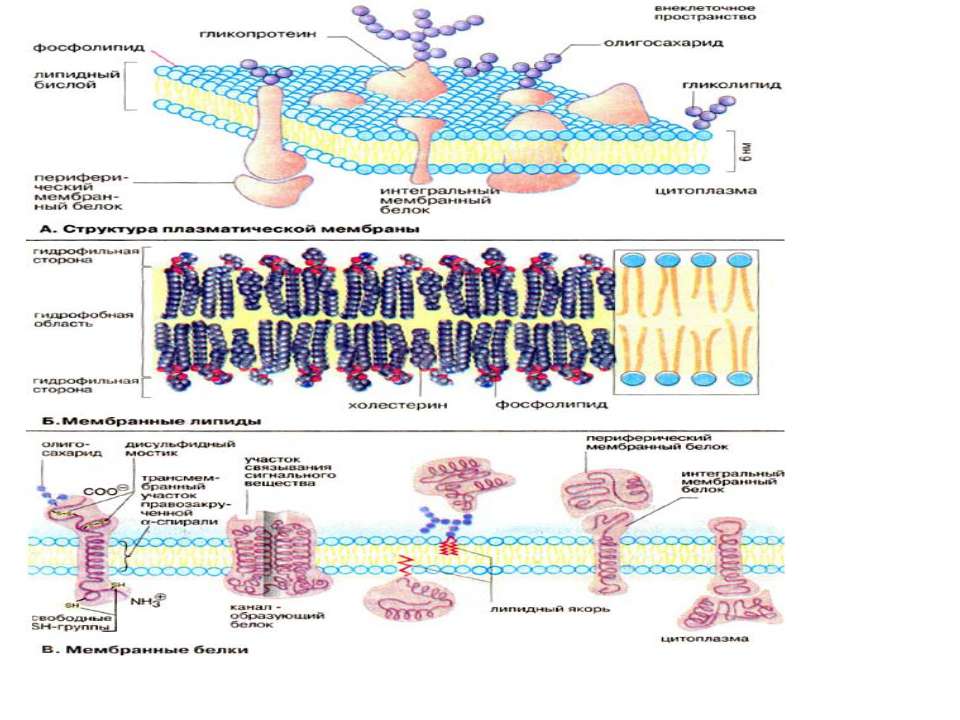

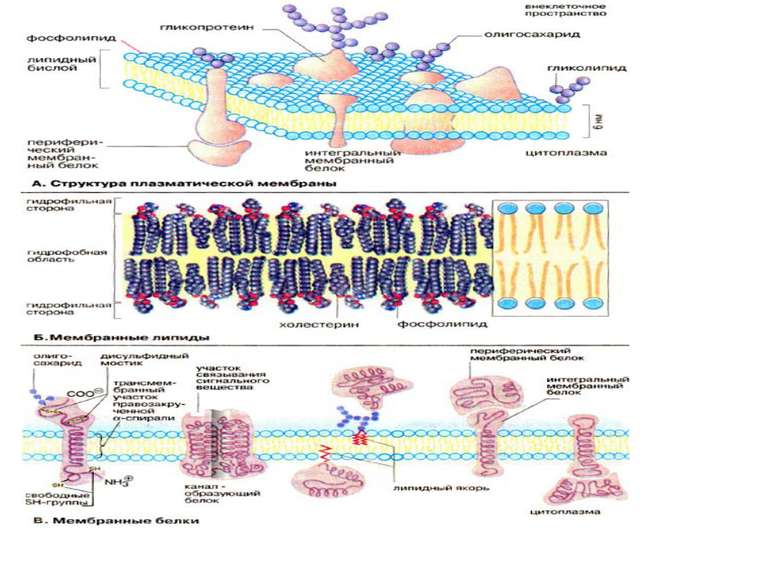
Всі біомембрани побудовані однаково; вони складаються із двох шарів ліпідних молекул біля 6 нм, в які вбудовані білки. Деякі мембрани містять, вуглеводи, зв’язані з ліпідами та білками. Співвідношення ліпіди : білки : вуглеводи є характерним для клітин або мембрани та суттєво варіює в залежності від типу клітин або мембран. Компоненти мембран утримуються нековалентними зв’язками, за рахунок чого вони мають властивість лише відносної рухливості тобто можуть дифундувать в межах ліпідного бішару. «Плинність» мембран залежить від ліпідного складу та температури мікрооточення. Із збільшенням вмісту ненасичених жирних кислот «плинність» збільшується, так як наявність подвійних зв’язків сприяє порушенню напівкристалічних мембранних структур. Рухливими є і мембранні білки. Якщо білки не закріплені в мембрані, вони «плавають» в ліпідному бішарі як в рідині, і тому існує таке поняття як рідинно-мозаїчна структура біомембрани. В той час як «дрейф» в площині мембрани відбувається достатньо легко, перехід білків із зовнішньої сторони мембрани на внутрішню («фліп-флоп») неможливий, а перехід ліпідів відбувається надзвичайно рідко. Для «перескоку» ліпідів необхідні спеціальні білки - транслокатори. Виключенням є холестерин, який може легко переходить з однієї сторони мембрани на іншу.

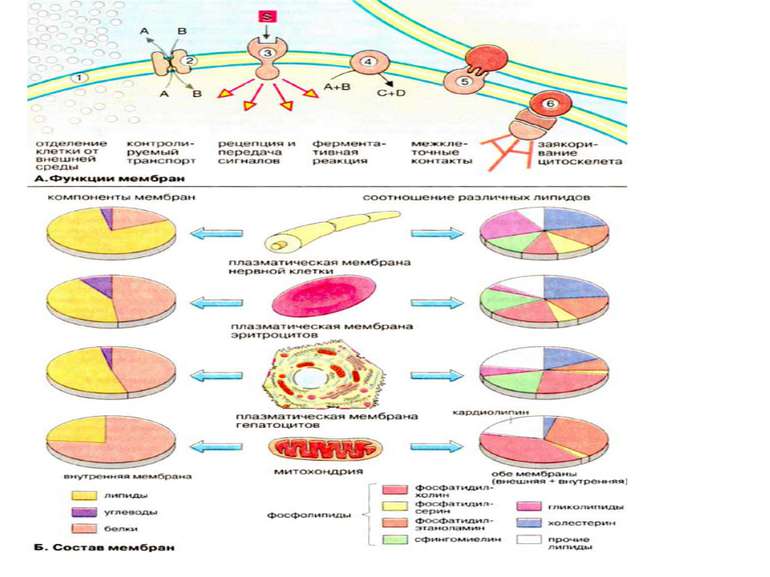
Мембранні ліпіди В мембранах містяться ліпіди трьох класів: фосфоліпіди, холестерин та гліколіпіди. Найбільш важлива группа, фосфоліпіди, включає фосфатітілхолін (лецитин), фосфатідилетаноламін, фосфатідилсерин, фосфатідилінозит та сфінгомієлин. Холестерин присутній у внутрішньоклітинних мембранах клітин (за виключенням внутрішньої мембрани мітохондрій). Гліколіпіди входять в склад багатьх мембран (наприклад, в зовнішній шар плазматичних мембран). В склад гліколіпідів входять вуглеводні функціональні групи, які орієнтуються в водну фазу. Ліпіди мембран являють собою амфіфільні молекули з полярною гідрофільною головкою та неполярним ліпофільним хвостом. В водному середовищі вони агрегують за рахунок гідрофобних взаємодій та вандерваальсових сил.

Мембранні білки Протеїни можуть зв’язуватись з мембраною різним шляхом. Інтегральні мембранні білки мають трансмембранні спіралізовані домени, які однократно або багатократно пересікають ліпідний бішар. Такі білки «прочно» зв’язані з ліпідним оточенням. Периферийні мембранні білки утримуються на мембрані за допомогою ліпідного «якоря» та зв’язані з іншими компонентами мембрани; найчастіше ассоційовані з інтегральними мембранними білками. У інтегральних мембранних білків фрагмент пептидного ланцюга, який пересікає ліпідний бішар, складається із 21-25 переважно гідрофобних амінокислот, які утворюють праву α-спіраль з 6 або 7 витками (трансмембранна спіраль). Периферийні білки легко відділяються від ліпідного шару при мінімальних обробках, інтегральні білки вбудовані і їх можна видалити лише при розрушенні мембрани. До деяких мембранних ліпідів та білків (гліколіпіди та глікопротеїни) приєднані олігосахаридні ланцюги. Плазматична мембрана може поповнювати свій склад (старі молекули можуть інтерналізуватись внутріклітинно та деградувати).

Класифікація мембранних білків може бути також проведена за їх функціональною приналежністю. Структурні білки: надають клітині та органелам певну форму надають мембрані та плазмолемі ту або іншу механічну функцію (еластичність, збудливість) забезпечують зв’язок мембрани з цитоскелетом, або у випадку ядерної мембрани з хромосомами Транспортні білки Проникність мембрани обумовлена ліпідним бішаром, хоча його можливості обмежені невеликою групою молекул, а основна група молекул переміщається через мембрану через транспортні білки, які поділяються на односторонні та двохсторонні. Створюються стійкі транспортні потоки певних речовин через мембрану, а також транспорт іонів призводить до змін трансмембанного потенціалу Мембранні рецептори - інтегральні компоненти мембрани, здатні активуватись зв’язуючись з специфічними лігандами. рецептори лектинів, які зв’язують певні моносахариди (наприклад конковалін А) рецептори кінцевих груп олігосахаридів рецептори сенсебілізованих лімфоїдних клітин, які зв’язують відповідні антигени (імуноглобулінові молекули) рецептори білкових гормонів та нейромедіаторів (інсулін) рецептори бактерільних токсинів (холерний токсин) адгезивні рецептори (інтегрини, селектини, кадгерини, імуноглодулінподібні молекули)
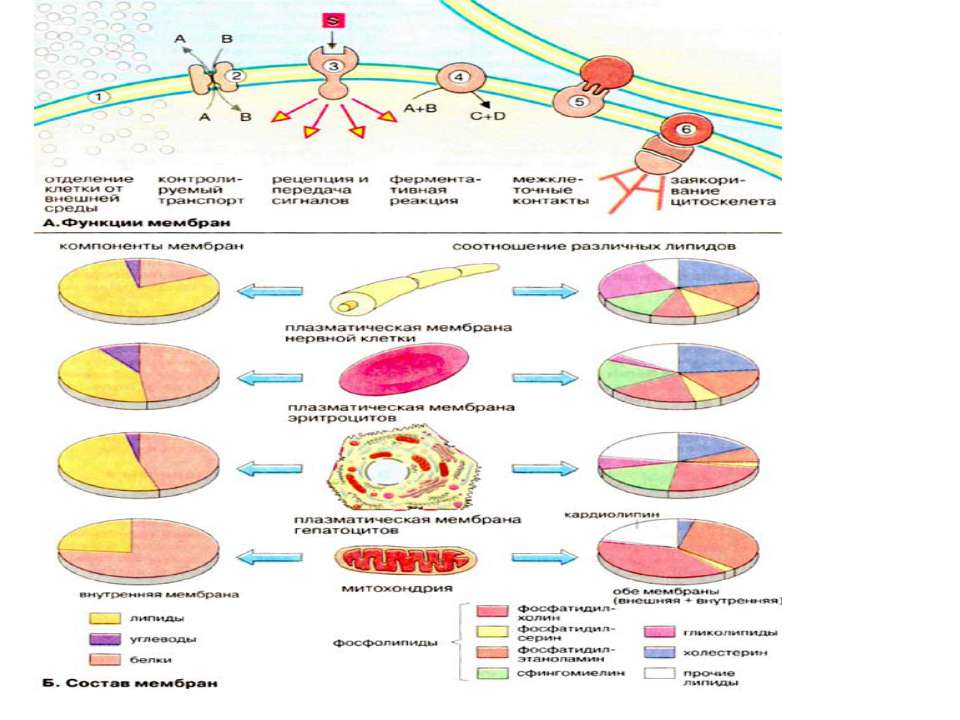

Функції біомембран Біомембрани виконують слідуючі функції: 1. Обмеження та обособлення клітин та органел. Обособлення клітин від міжклітинного середовища забезпечується плазматичною мембраною, яка захищає клітину від механічного та хімічного подразнення. Плазматична мембрана забезпечує також збереження різниці концентрацій метаболітів та неорганічних іонів між внутріклітинним та зовнішнім середовищем. 2. Контрольований транспорт метаболітів та іонів визначає внутрішнє середовище, що є суттєвим для гомеостазу, тобто підтримання постійної концентрації метаболітів та неорганічних іонів, а також інших фізіологічних параметрів. Регульваний селективний транспорт метаболітів та неорганічних іонів через пори та опосередковано білками-переносниками стає можливим із-за обособлення клітин та органел за допомогою мембранних систем. 3. Сприйняття позаклітинних сигналів, їх внутріклітинна передача та ініціація сигналів. 4. Ферментативний каталіз. В мембранах на межі між ліпідною та водною фазами локалізовані ферменти. Якраз тут відбуваються реакції із неполярними субстратами Прикладом може слугувати біосинтез ліпідів та метаболізм неполярних ксенобіотиків. В мембранах локалізовані найбільш важливі реакції енергетичного обміну, такі, як окислювальне фосфорилювання та фотосинтез (рослинні клітини) 5. Контактна взаємодія з міжклітинним матриксом та взаємодія з іншими клітинами при при злитті клітин та утворенні тканин. 6. Заякорення цитоскелету, яке забезпечує підтримання форми клітин та органел та клітинної рухливості.

Слід підкреслити, що в фізіологічних умовах ліпіди, які утворюють мембрану, знаходяться переважно в рідко-кристалічному виді. Це означає, що кожна молекула внутрі свого моношару має високу рухливість, але перехід молекул із одного ліпідного моношару в інший спостерігається дуже рідко. Зміни мікров’язкості ліпідного шару можуть бути результатом зміни співвідношення різних хімічних класів ліпідів в мембрані. Багато рецепторів здатні рухатись в ліпідному шарі, проте існують механізми, які обмежують цю рухливість: - заякорення рецепторів ззовні, тобто прикріплення їх до якоїсь структури ззовні від мембрани, заякорення рецепторів внутрі клітини до кортикальних структур, що розміщені під мембраною агрегація рецепторів в площині мембрани.

Свіжеотримані первинні клітини і висаджені в культуру мають сферичну форму. Механізми, які сприяють розпластання клітин на підложках слідуючі: Початкове прикріплення – клітина прикріплюється нижньою поверхнею до підложки, проте ніяких виростів ще нема, лише в області контакту мембрана потовщена, але кортикальний шар ще не змінений. Утворення ламелярної цитоплазми – відсутні крупні органели і існують маса точок прикріплення до підложки. Стадія поляризації. Різні типи псевдоподій, які утворюються при розпластанні клітин на поверхні мають ряд загальних властивостей: -фокальні контакти між клітиною і підложкою утворююються на поверхні псевдоподій, але не на поверхні інших частин клітини -в цитоплазмі псевдоподій утворюється центробіжне натягнення, інертні частинки, які попадають на поверхню псевдоподій і ламелярної цитоплазми, прикріплююються до цієї поверхні і переміщуються по ній центробіжно -розвиток натягнення, в прикріпленій псевдоподії зберігається натягнення, бо при їх розрушенні псевдоподії та ламілярна цитоплазма моментально скорочується очистка від склеєних рецепторів, які після приєднання лігандів агрегують відбувається слідуючим чином: спершу очищуються кінці псевдоподій по центробіжному параметру (від кінця псевдоподій до центру).

механізм псевдоподіальних реакцій може включати в себе декілька стадій: - початковий етап- це іммобілізація группи рецепторів мембрани - утворення такої групи рецепторів може індукувати кортикальні зміни, що приводять до появи псевдоподій; -якщо утворена група імобілізованих рецепторів знаходиться на поверхні псевдоподій, то вони індукують іншу групу кортикальних змін, що приводять до заякорення цієї групи рецепторів внутрі і до їх з’єднання з актиновими мікрофіламентами, в результаті в місті контакту розвивається цетробіжний натяг в залежності від умов реакції прикріплення або натягу процес закінчується або прикріпленням і утворенням контакту, або центробіжним переміщенням імобілізованих рецепторів. Для трансформованих клітин характерне змінене формування мікровиростів та погіршене розпластання трансформованих клітин на поверхні. Ці характеристики можуть бути як причиною так і наслідком трансформованності клітин

Поляризовані трансформовані клітини також, як і нормальні здатні до активного переміщення по підложці, однак деякі статистичні характеристики цього переміщення можуть змінюватись при трансформації. Трансформовані клітини більш часто міняють напрямки при міграції (це показано мікрокінематографічними зйомками, а також по переміщенню клітин на підложці з колоїдним золотом). Такі зміни руху трансформованих клітин є вторинною ознакою дефектного прикріплення клітин до підложки: погано прикріплені вузькі активні місця легше відриваються від підложки, чим широкі добре прикріплені місця нормальних клітин. Стикання активного краю клітини, що рухається з краєм іншої клітини може призводити до трьох результатів: зупинці руху, підповзанню, або наповзанню клітин одна на іншу. При прямому спостереженні за поведінкою при русі трансформованих клітин було виявлено, що для трансформованих клітин характерне також пригнічення руху (зупинка, відсутність наповзання одна на іншу). Котнактне гальмування у трансформованих клітин частіше чим у нормальних супроводжується контактним паралічем та ретракцією. Ретракція у трансформованих клітин виражена більш сильніше, чим у нормальних і приводить до повного відриву від підложки та скороченню цитоплазматичних відростків, на яких розміщений активний край.

З точки зору псевдоподіальних реакцій різні культури трансформованих клітин можуть мати значне морфологічне різноманіття. Так, при дії на нетрансформовані клітин одним і тим же агентом можуть виникати різні трансформовані клони, які можуть мати різну морфологію, наприклад: -Одношарові або багатошарові культури, які складаються із погано розпластаних циліндричних клітин (зокрема фібробластів), в яких сусідні клітини паралельні одна іншій -Багатошарові культури, які складаються із погано розпластаних циліндричних полігональних або зірчатих клітин -Багатошарові клітини, які складаються в основному із округлих клітин -Одношарові культури, які складаються в основному із майже сферичних клітин, які сидять на підложці.

МІЖКЛІТИННІ КОНТÁКТИ — з’єднання, що встановлюються між сусідніми клітинами у складі тканин та органів багатоклітинних організмів. Вони можуть бути різними як за формою, структурою, так і за забезпеченням контактів між клітинами. За різними джерелами наукової інформації класифікація М.к. може бути різною, але найбільш поширеним є їх поділ на прості та складні. Просте неспеціалізоване міжклітинне з’єднання — це контакт плазмалем клітини на відстані 10–20 нм, при якому взаємодіють шари глікокаліксу обох клітин. Контактуючі поверхні клітин можуть бути паралельними чи утворювати взаємні вростання, коли вирости плазмалеми і цитоплазми однієї клітини занурюються у відповідні заглибини сусідньої клітини. Такий тип контакту носить назву зубчастого або пальцеподібного.

Складні контакти, у свою чергу, можуть бути щільними замикальними (ізолюючими), заякоряючими (зчеплювальними) та комунікаційними. Щільний замикаючий контакт характерний для клітин епітеліальної вистілки травного тракту і епітелію залоз. При формуванні щільного контакту зовнішні шари мембран на окремих ділянках максимально зближуються, внаслідок чого стають непроникними для макромолекул та іонів; також спостерігається утворення одного суцільного пласта, який пояском оточує апікальні ділянки клітин. Заякоряючі, або зчеплювальні контакти утворюються за участю фібрилярних елементів цитоскелета. До цих міжклітинних з’єднань належать зчеплювальні стрічки, фокальні контакти та бляшки зчеплення, які пов’язуються з актиновими мікрофіламентами всередині клітини, а також десмосоми та напівдесмосоми, які з’єднуються з іншими елементами цитоскелета — проміжними філаментами. Зміцнення контакту між клітинами досягається шляхом формування десмосом — утворень цитоплазми двох сусідніх клітин, кожна з яких формує товсту пластинку прикріплення діаметром до 0,5 мкм. Між пластинками знаходиться міжклітинна щілина шириною 25–30 нм, заповнена електронно-щільною речовиною, утвореною молекулами інтегральних глікопротеїнів — десмоглеїнів. З боку гіалоплазми в зоні десмосоми розташовується електронно-щільний шар білка — десмоплакіну, в який вплітаються проміжні елементи цитоскелета.

Десмосоми є характерними контактами епітеліальних, ендотеліальних клітин, кардіоміоцитів та інших клітин. На відміну від десмосоми, що складається з двох пластинок прикріплення, напівдесмосома має лише одну таку пластинку і утворюється в місцях контакту епітеліальних клітин із базальною мембраною. На відміну від щільного контакту, всі типи зчеплювальних контактів є проникними для водних розчинів і не відіграють ніякої ролі в обмеженні дифузії. Комунікаційні, чи щілинні, міжклітинні контакти є функціональними зв’язками між клітинами. Через щілинні з’єднання клітини здійснюється прямий обмін хімічними речовинами між клітинами. До них належать нексуси та різні групи синапсів. Нексус — це спеціалізований клітинний контакт, який відзначається безпосереднім хімічним зв’язком між цитоплазмами клітин, коли плазмалеми сусідніх клітин зближені до відстані 2–3 нм і пронизані особливими часточками — конексонами, кожна з яких складається з 6 субодиниць із циліндричним каналом по центру. У складі різних щілинних контактів нараховується від кількох одиниць до декількох тисяч конексонів. Через конексони утворюються наскрізні канали, які сполучають між собою внутрішні середовища контактуючих клітин. Нексуси містяться переважно в серцевій м’язовій тканині й забезпечують тісний метаболічний зв’язок між цитоплазмами контактуючих кардіоміоцитів.

Синапси — це спеціалізовані контакти між нейронами або між нейронами та м’язами, які служать для передачі збудження або гальмування в один бік від однієї клітини до іншої. Синапси утворюються на термінальних ділянках відростків нейронів — дендритів та аксонів. Один нейрон за посередництвом синапсів може з’єднуватися з 10 000 нейронів. Міжнейронні синапси підрозділяються на електричні та хімічні. Мембрани клітин, що утворюють синапс, розділені міжклітинним простором — синаптичною щілиною шириною 20–30 нм, через яку передається інформація у вигляді медіаторів. Таким чином, різні типи міжклітинних контактів пристосовані до особливих функцій певних клітин.

Щільні контакти Цей тип міжклітинних з'єднань широко представлений в епітеліальній тканині. Він вносить суттєвий вклад у функціональну асиметрію епітеліоцитів і впливає на характер вибіркової проникності, зумовлюючи структурну асиметрію плазматичної мембрани.Сформовані щільні контакти лежать в основі структурно-функціональної поляризації багатьох епітеліїв. Мембрана, обернена до зовнішнього середовища – апікальна мембрана – активно транспортує певні молекули до клітини. Друга поверхня клітин (базолатеральна) складається з двох компонентів: латеральної мембрани – поверхні, що стикається з сусідніми клітинами, і базальної мембрани – поверхні контактування з позаклітинною рідиною та позаклітинним матриксом. Це означає, що певні клітинні білки (наприклад, переносники вуглеводів і амінокислот, іонні насоси тощо) розташовані або лише на апікальній, або на базолатеральній поверхні клітин. Крім того, аналіз взаємодії епітеліальних клітин з різними мембранними білками вірусів, виявив їх вибіркове зв'язування або з апікальною, або з базолатеральною плазматичними мембранами. Відбруньковування вірусних часток також здійснюється від різних поверхонь інфікованих клітин. Це є можливим лише за умови підтримання асиметрії плазматичних мембран, що й забезпечується щільними контактами, які перешкоджають дифузії мембранних білків між апі кальною та базолатеральною поверхнями мембрани й щільно змикають сусідні клітини, попереджаючи проходження розчинних молекул між ними

Інтегрини беруть участь в утворенні трьох типів адгезивних контактів: "клітина-клітина", "клітина-матрикс" і "клітина-розчинний фактор". Інтегрини – гетеродимерні білки: вони складаються з двох нековалентно зв'язаних субодиниць . Кожна субодиниця має цитоплазматичний, трансмембранний і позаклітинний домени. Цитоплазматичний домен взаємодіє з цитоскелетом клітини, позаклітинний – зв'язується з компонентами позаклітинного матриксу. Така взаємодія забезпечує двобічну передачу сигналу, наприклад, актинові філаменти цитоскелета клітин здатні змінювати орієнтацію молекул фібронектину, що секретуються, у позаклітинному матриксі.Інтегрини виявляють активність за наявності іонів кальцію та магнію і беруть участь у передачі сигналів, які регулюють експресію генів і проліферацію

Кадгерини. За їхньою участю формуються гомофільні адгезивні контакти які забезпечуються взаємодією подібних молекул адгезії (в контактах типу "клітина – клітина") за наявності іонів кальцію. До цього класу відносять понад 20 інтегральних високомолекулярних білків. Так, Е-кадгерини експресуються на поверхні епітеліальних клітин у складі проміжних міжклітинних контактів і клітин концептусу до імплантації (увоморулін), а потім на пізніших стадіях розвитку (наприклад, у клітинах нейроектодерми). Вони зменшують рухливість клітин пухлин епітеліальних тканин. N-кадгерин присутній у нейронах, скелетному та серцевому м'язах, клітинах кришталика (бере участь у процесі подовження відростка нейрона). М-кадгеринз'являється в мітогенезі скелетного м'яза на етапі формування трубочок. Р-кадгерин виявлений у кератиноцитах і клітинах плаценти.Другий клас молекул адгезіїскладають імуноглобуліни, які здійснюють як гомофільні, так і гетерофільні адгезивні контакти, в яких беруть участь адгезивні молекули різних класів (це контакти типу "клітина-клітина" і "клітина-субстрат").
Схожі презентації






Категорії











