Презентація на тему:
Мінливість у людини як властивість життя і генетичне явище
Завантажити презентацію




– ступінь фенотипового прояву одного...")













 Жидкостная цитометрия, обычно используемая дл...")





 Следующей трансформацией цитогенети...")
")

, температура...")


 реактивація Уейгла (індукція додаткових репаративних...")
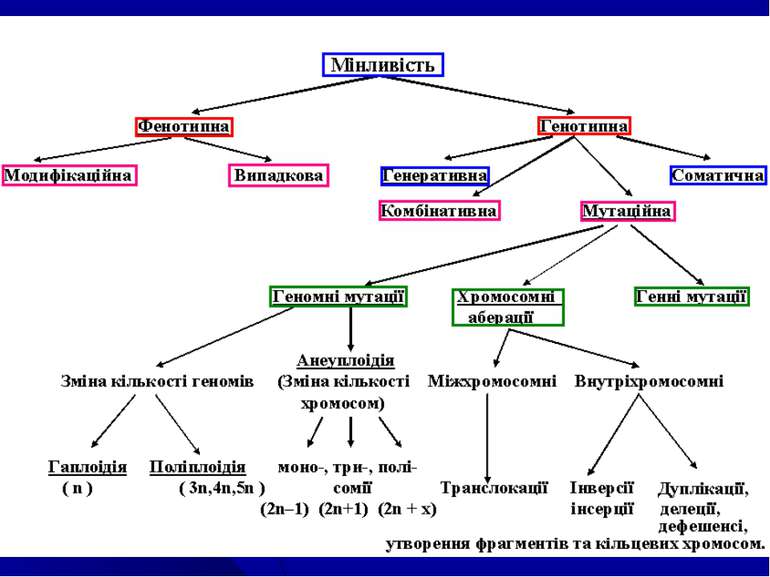
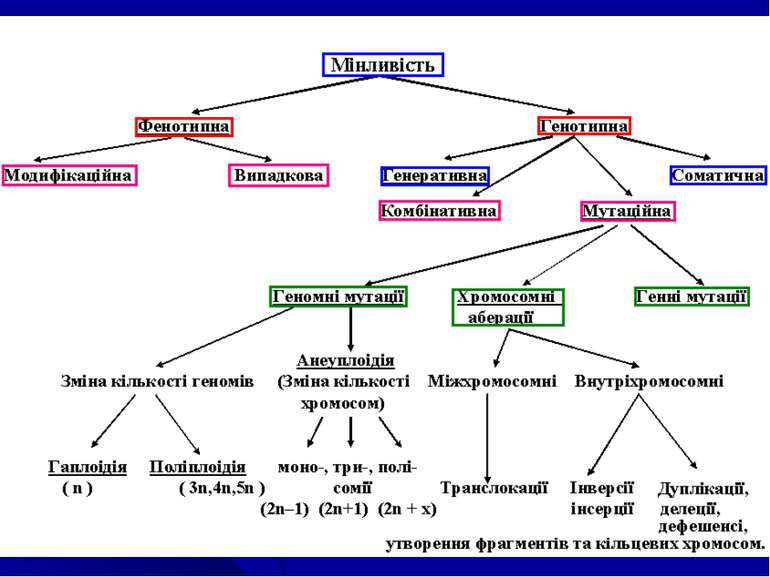
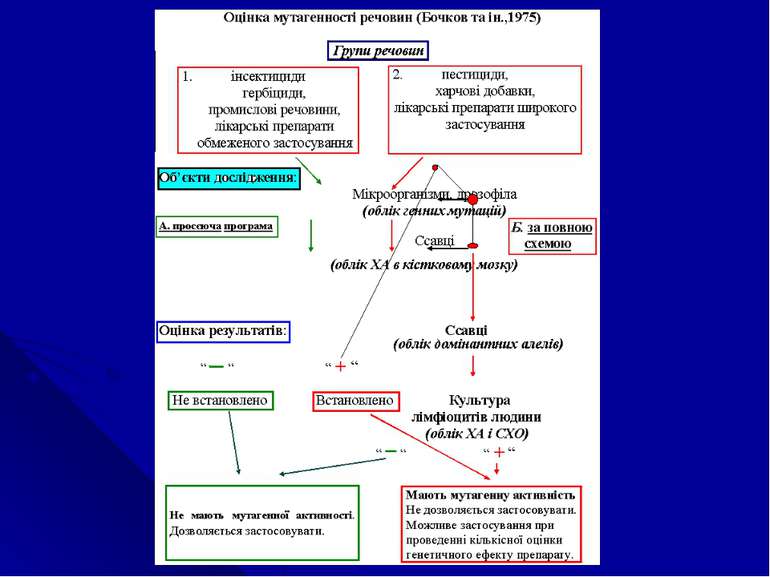
 1. Види...")
 організмів взагалі характеризуються певним циклом гомо...")


Мінливість у людини як властивість життя і генетичне явище
Завантажити презентаціюПрезентація по слайдам:

Мінливість у людини як властивість життя і генетичне явище.

Питання теми: Мінливість та її форми. Модифікаційна мінливість: норма реакції генетичне детермінованих ознак, експресивність та пенетрантність, фенокопії, адаптивний характер модифікацій. Комбінативна мінливість – основа для генетипової та фенотипової різноманітності людства. Мутаційна мінливість. Класифікація мутацій: Мутагенні фактори. Антимутагенні механізми на різних рівнях організації життя. Репарація. Закон гомологічних рядів спадкових форм мінливості. Оцінка мутагенності зовнішніх факторів.

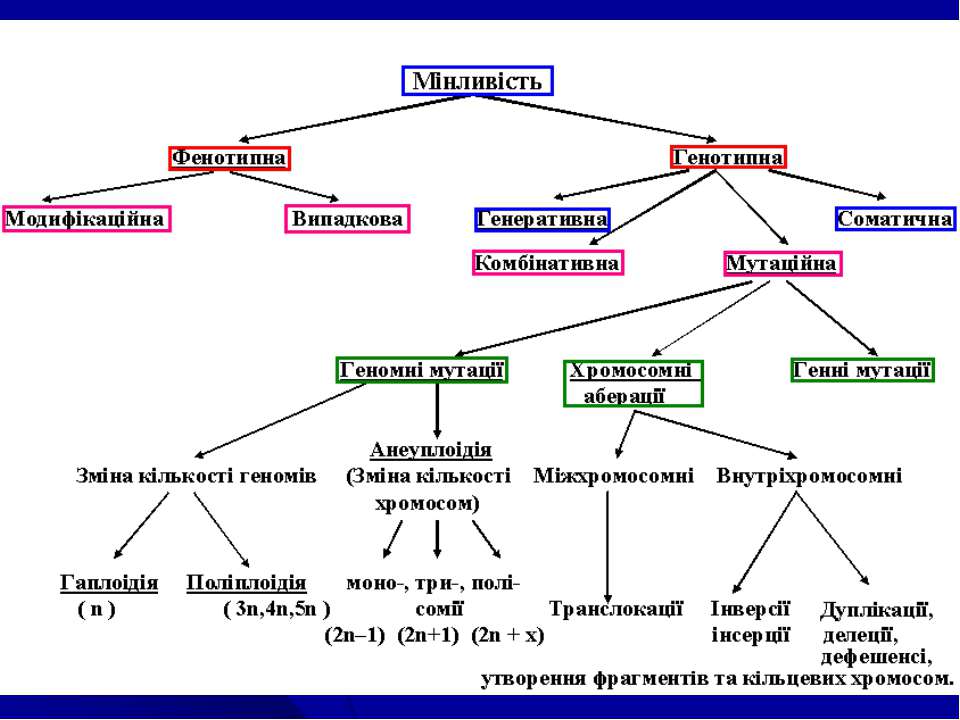
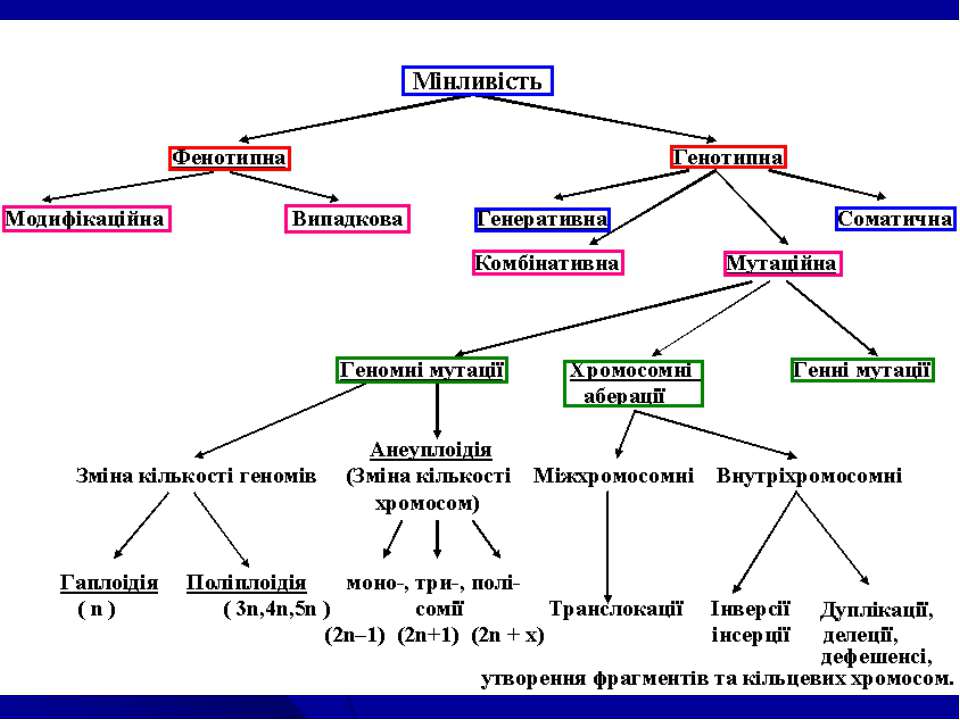
Основні поняття: Мінливість – властивість організмів у межах виду існувати в різних варіантах ознак і властивостей, - властивість організмів набувати в процесі індивідуального розвитку нові властивості на ознаки. Виділяють М.: кореляційну, визначену, невизначену, онтогенетичну (Ч.Дарвін). З генетичної точки зору: фенотипна та генотипна. Реалізується на клітинному та організменому рівнях в процесі їх розвитку. Фенотипна М. – зміна(и) фенотипу під дією зовнішніх факторів, умов існування виду, які не викликані порушеннями генотипу. Генотипна М. – зміна(и) фенотипу внаслідок перекомбінування або мутації спадкового матеріалу на різних рівнях його організації (каріотипу, хромосом, генотипу). Норма реакції – діапазон модифікаційної мінливості генетично детермінованих ознак в різних умовах існування виду. Норма реакції успадковується (для одних ознак – широка, для інших – вузька).
– ступінь фенотипового прояву одного...")
Експресивність (Тімофєєв-Рисовський,1927)– ступінь фенотипового прояву одного і того ж алелю гену у різних особин. При відсутності мінливості ознаки, яка контролюється даним алелем гену Е. – постійна, у випадках різних варіант ознаки Е називають варіабільною. Причини: а) вплив різних умов середовища, б) модифікуюча дія інших генів в однакових умовах середовища, в) залежность від комбінації певних генів в генотипі. Пенетрантність (Тімофєєв-Рисовський,1927) – частота фенотипового прояву алелю гену у різних особин родинної групи організмів. Виділяють повну П. (алель проявляється у всіх особин, які мають його в генотипі) і неповну П. (алель фенотипове не проявляється у частини особин, які мають її в генотипі). Фенокопії (Р. Гольдшмідт, 1935) – неспадкові зміни фенотипу (модифікації), які нагадують певні відомі зміни фенотипу при мутаціях. Ф. – результат дії фізичних або хімічних факторів на генетично нормальний організм в процесі його індивідуального розвитку в певні, фенокритичні стадії (періоди) онтогенезу.

Норма реакції


Норма реакції

V— варіанти ознаки, Р — частота варіантів ознак, Мо — мода, або найчастіше значення ознаки, lim — межі модифікаційної мінливості ознаки (норма реакції) - Модифікаційна крива:

Адаптивний характер модифікацій

Зміни активності генів, Зміни процессів транскрипції та трансляції, Зміни проліферації клітин, диференціації…

Хромосомний аналіз

Впервые хромосомы человека наблюдали Flemming и Arnold в 1880-х годах Цитогенетика человека начинает свой отсчет с открытия, сделанного в 1956 году, когда Tjio и Levan добавив воду к суспензии митотических клеток человека перед их фиксацией на стекле, смогли отделить хромосомы друг от друга и сосчитать их количество. Независимо от их исследования, в том же году, число хромосом человека - 46, было установлено Ford и Hamerton

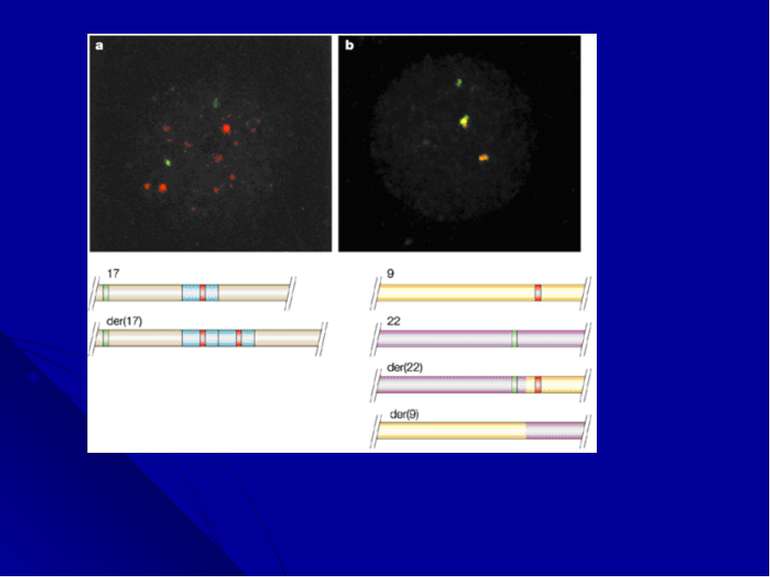
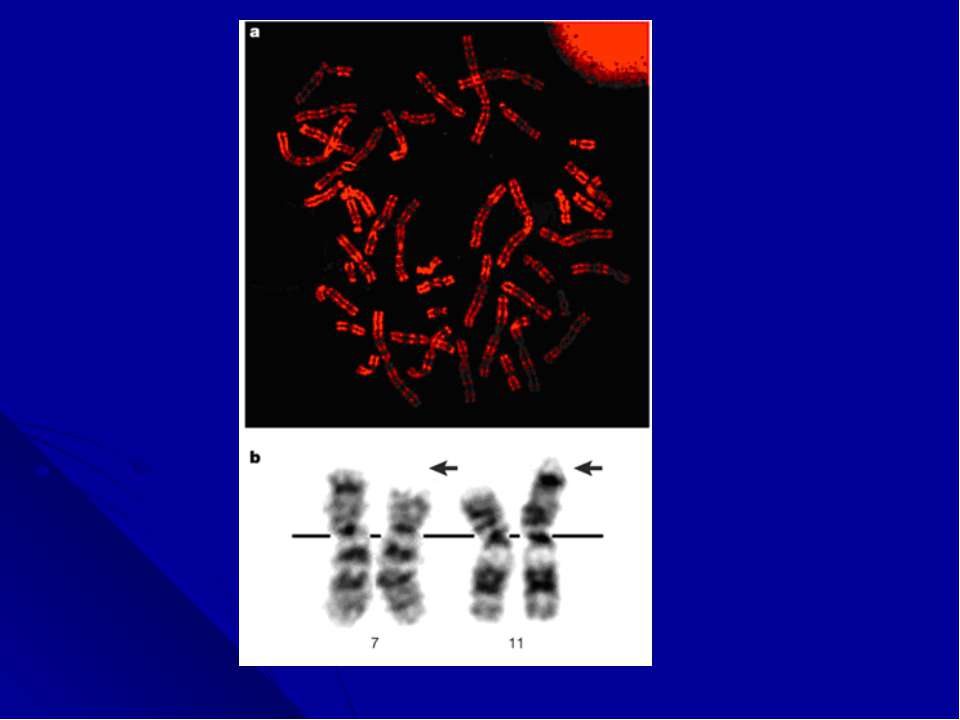
«Филадельфийская» хромосома «Филадельфийскую» хромосому как причину хронического миелоидного лейкоза (ХМЛ) идентифицировали в 1960 году. Через 30 лет, благодаря появлению молекулярного анализа, Janet Rowley обнаружила, что она является продуктом транслокации между хромосомами 9 и 22 , а в 1985 году, в месте слияния этих хромосом, был идентифицирован новый гибридный ген, состоящий из двух генов (BCR и ABL). Дальнейшие исследования показали, что активация этих генов тирозинкиназой лежит в основе патогенетических механизмов развития фенотипа болезни Это открытие способствовало созданию лекарства, которое блокирует функцию белка BCR-ABL и с успехом применяется для лечения больных ХМЛ.

Изучение первых препаратов хромосом человека в начале 1960-х способствовало и другим открытиям в генетике человека. Lejeune в 1963 году описал первый делеционный синдром – cri du chat - потерю порции короткого плеча хромосомы 5 у больных с выраженной задержкой умственного развития и плачем, похожим на крик кошки

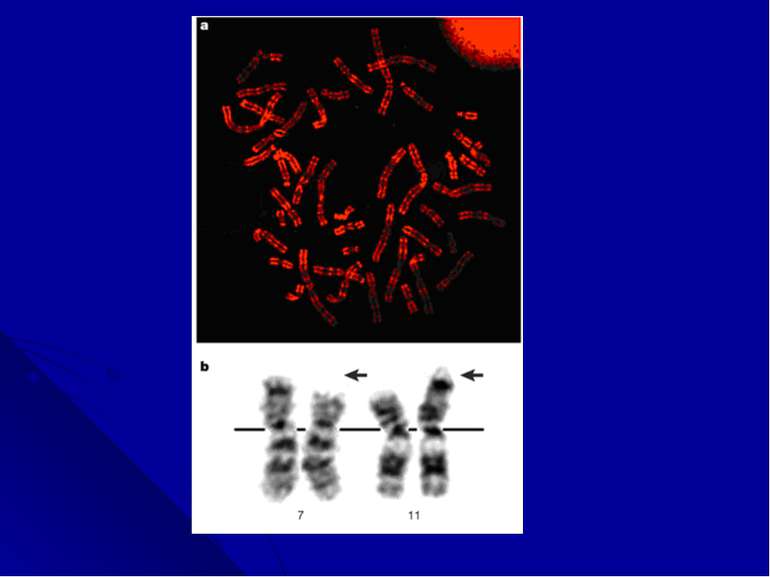
Дифференциальное окрашивание Цитогенетические исследования получили новое развитие в конце 1960-х годов благодаря работам Torbjorn Caspersson, который разработал дифференциальное окрашивание, дававшие паттерн из темных и светлых полос по всей длине каждой хромосомы. В прометафазе, когда хромосомы находятся на очень ранней стадии конденсации, можно было насчитать до 2000 полос, в других случаях - 400-800 полос. Эта окраска позволила цитогенетикам легко идентифицировать хромосомы, выявлять делеции, инверсии, инсерции, транслокации и другие более сложные перестройки.

Следующим шагом после идентификации аномальной хромосомы является определение точки транслокации или границы делеции на молекулярной карте относительно генов. Для этого используются три подхода: технология гибридных соматических клеток, активизированная флюоресценцией сортировка хромосом (FACS) и FISH-метод. Эти методы обеспечивают грубое картирование, перед более детальным молекулярным анализом и секвенированием.

Гібридизація соматичних клітин
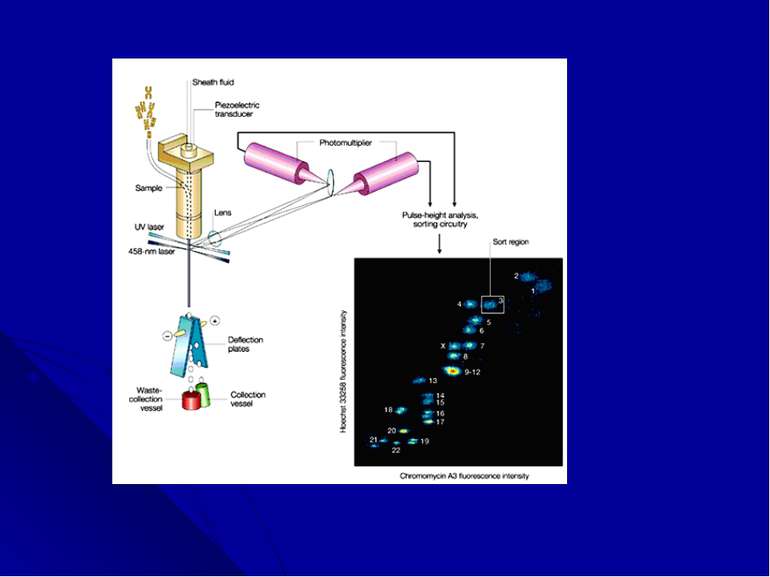
 Жидкостная цитометрия, обычно используемая дл...")
Жидкостная цитометрия (сортинг) Жидкостная цитометрия, обычно используемая для анализа и разделения клеток, в 1979 году группой исследователей (лаборатория в Калифорнии) была адаптирована для количественного анализа и сортировки хромосом. Этот метод позволяет отобрать все, кроме четырех хромосом человека [9-12] и получить информацию о вариабельности их размеров. Жидкостной сортинг лежал в основе создания библиотеки клонов хромосом-специфичной ДНК , которая используется для построения физической карты генома.


FISH-метод Следующим революционным достижением цитогенетики был метод FISH, который обеспечил прямую связь между микроскопом и секвенированием. Эта техника позволила увидеть хромосомную локализацию специфических последовательностей ДНК с помощью микроскопа

Прогрессивность методики заключалась в замене радиоактивного мечения ДНК и РНК проб, применяемого с 1969 года, на флюоресцентное. Флюоресцентные красители являются мягкими и простыми в употреблении, могут храниться неопределенно долго, дают высокое разрешение, и позволяют одновременно исследовать несколько последовательностей ДНК.

Техника FISH продолжала совершенствоваться, и сегодня локализация сегментов длиною10 kb считается рутинной, а - 1 kb вполне достижимой. Одним из последних достижений FISH является использование проб, состоящих из пептидов и нуклеиновых кислот, которые позволяют по интенсивности сигнала определять количество комплементарных последовательностей. Хорошей иллюстрацией является изучение динамики теломер в клетках с использованием TTAGGG-специфических проб к концам хромосом. Использование COD-FISH позволяет определять инверсии и сестринские хроматидные обмены

Что еще более важно, FISH сделала доступными для цитогенетического анализа ядра не делящихся клеток. Идентификация хромосом осуществляется простым подсчетом сигналов в каждом ядре. Изменение числа сигналов свидетельствует о произошедшей делеции или амплификации гена. Так дупликация длиной 1Mb, которая является причиной синдрома Шарко-Мари-Тус [61], может быть определена интерфазной FISH. Позиционные сдвиги сигналов выявляют структурные перестройки, такие как транслокации и инверсии
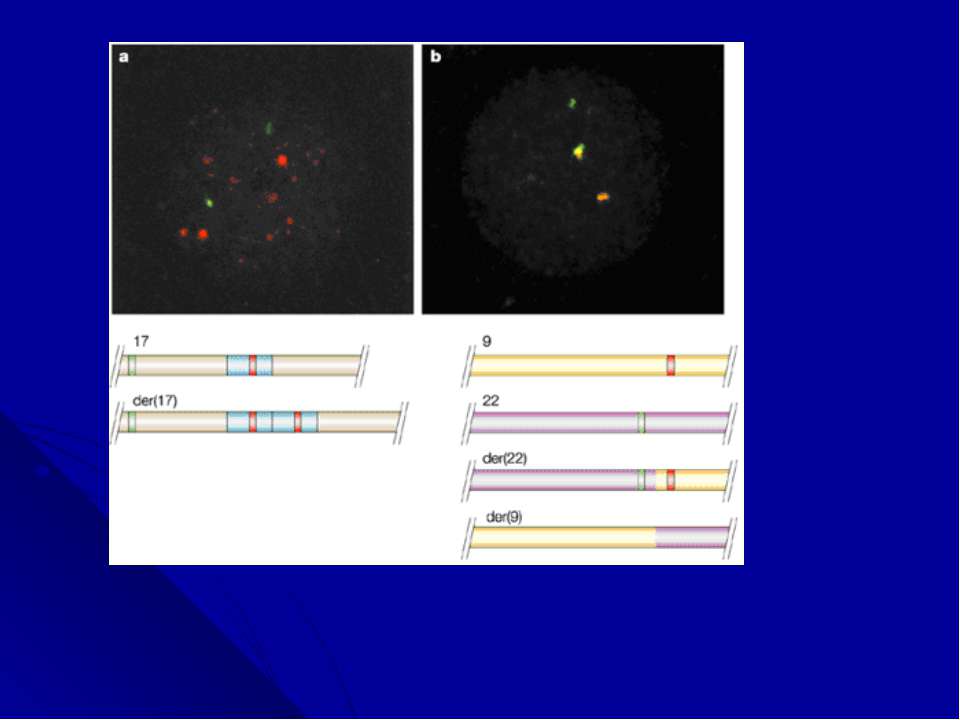

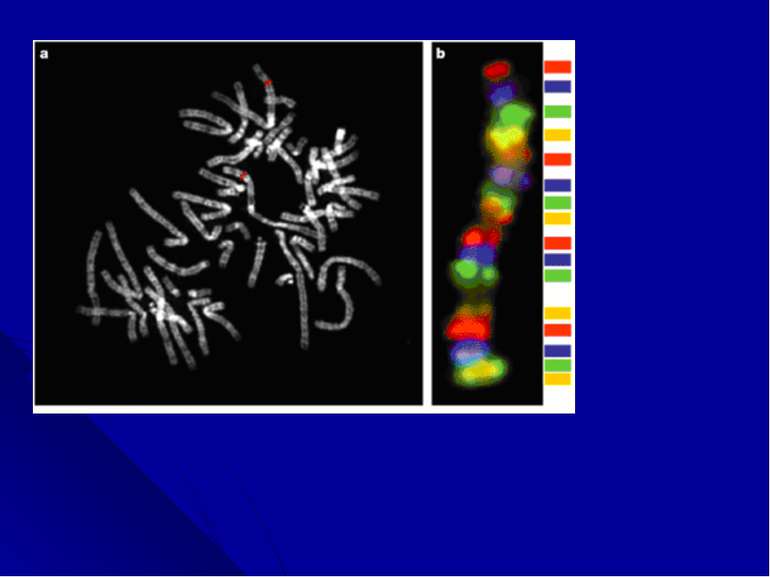
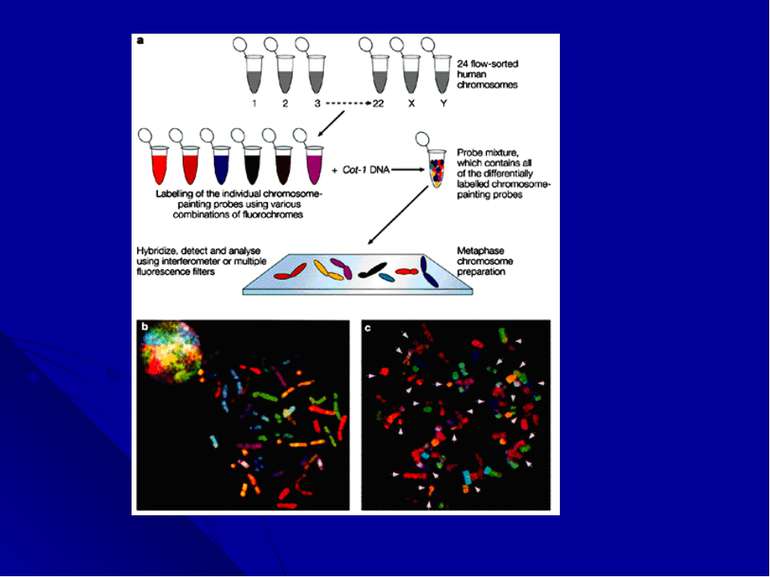
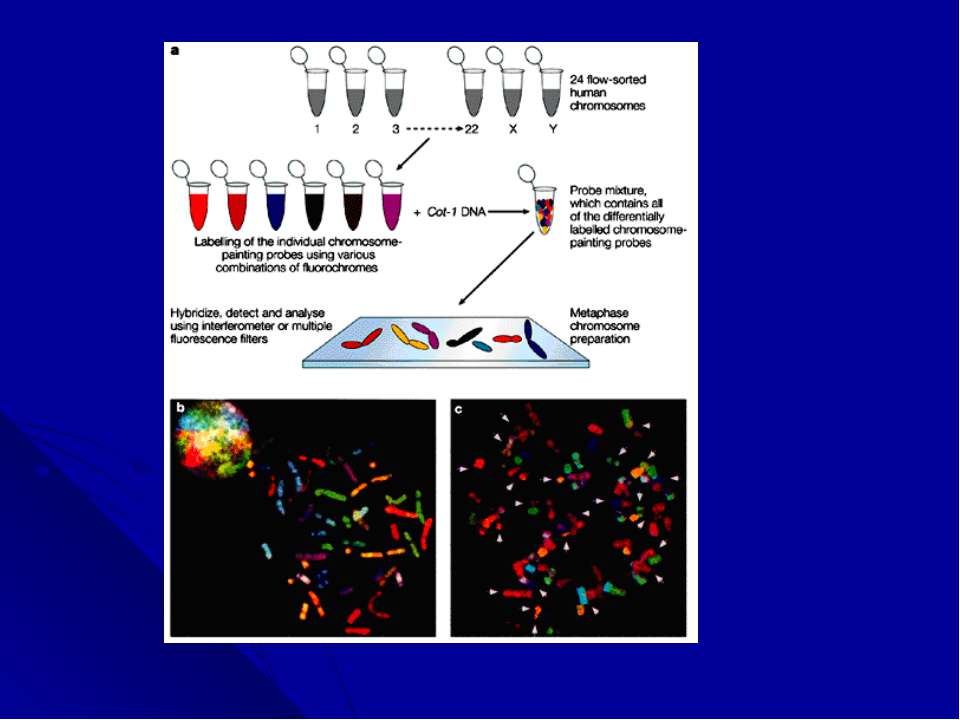
Многоцветное окрашивание хромосом. Последним достижением цитогенетических технологий является анализ многоцветно окрашенных хромосом. Новая методика, получившая название спектрального кариотипирования (SKY) или мультиплексной FISH (M-FISH), появилась в результате трех новшеств: 1) были созданы хромосом-специфические «краски»: коллекции последовательностей, выделенных из каждой хромосомы; 2)за счет сочетания флюорохромов, для каждой хромосомы получена своя 24-х цветовая комбинация; 3)для многоцветного анализа усовершенствована техника оптической микроскопии, системы фильтрации и отображения, позволяющая автоматически классифицировать каждый хромосомный сегмент.
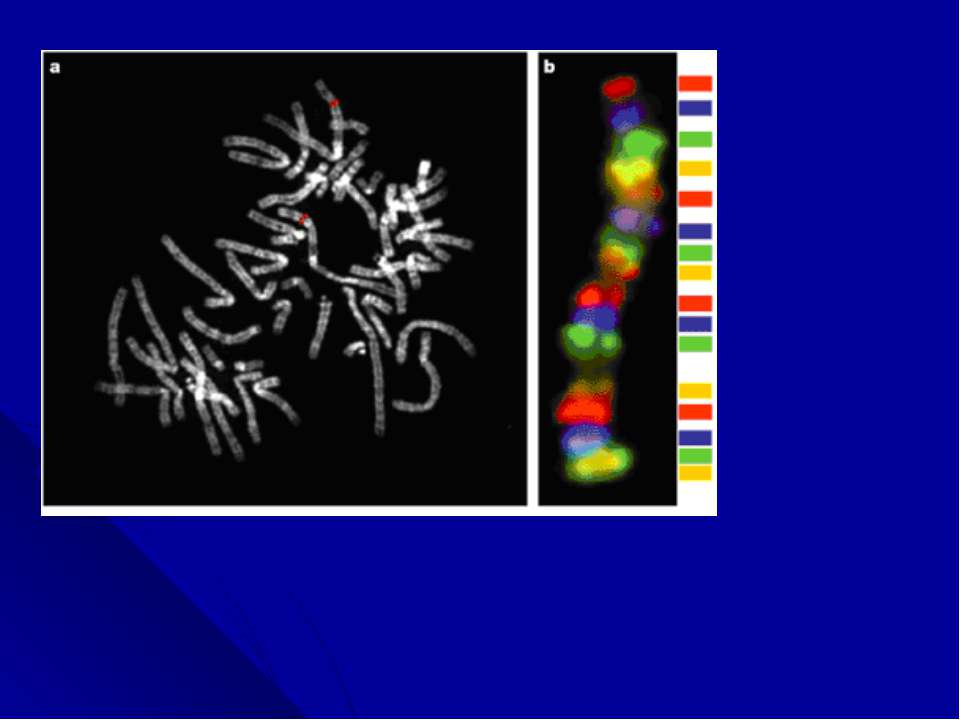
 Следующей трансформацией цитогенети...")
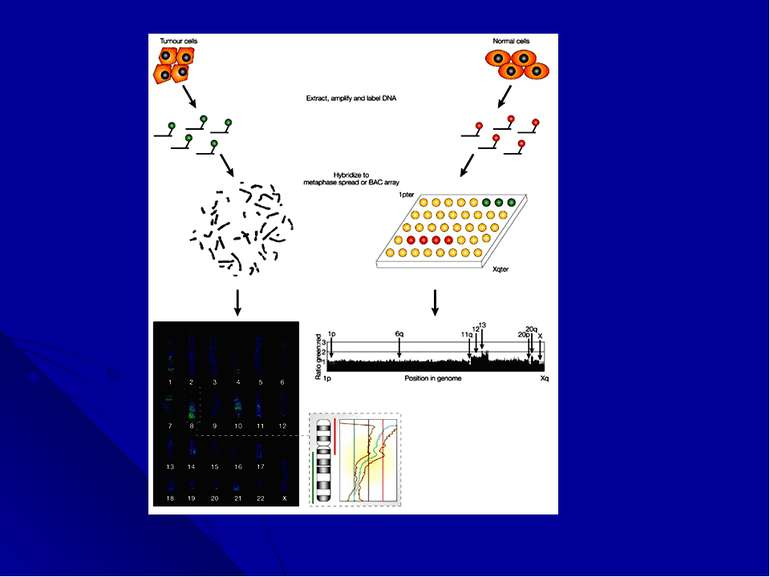
Сравнительная геномная гибридизация (CGH) Следующей трансформацией цитогенетики является реализация широкого сканирования генома на потерю или увеличение хромосомного материала без прямого осмотра хромосом -- сравнительная геномная гибридизация (CGH) В настоящее время цитогенетика вынашивает идею множественной-CGH

")
Геномні мутації. Анеуплоїдія Каріотип хворого при синдромі Едварса (трисомія 18)

Хромосомні аберації
, температура...")
Мутагенні фактори Фізичні: Ионізуюче випромінювання; УФ (260 нм), температура Хімічні: Солі важких металлів; Пестициди, промислові речовини, харчові добавки, ліки… Біологічні: - вируси, - бактерии, найпростіші, гельмінти; - трансгенні продукти !

Порушення ДНК Відбувається під дією зовнішніх факторів та спонтанно: апурінізація ( - пурінові основи). Кожна клітина за добу втрачає близько 5000 пурінів, внаслідок чого утворюється АР-сайт ( залишок дезоксирибози). Це відбувається спонтанно, дезамінування основ ( - NH2). При дезамінуванні виникають генні мутації ( Ц У( на добу 100 випадків на клітину), А гіпоксантин, Г ксантин. Продукти дезамінування визначаються репаративними системами клітини. Більшість канцерогенів – алкілірують (додають до азотистих основ групи -СН3 або –С2Н5 ). Під дією ультрафіолету відбувається утворення піримідинових димерів (Т-Т, Ц-Ц).

Репарація Пряма реактивація - ферментативне вирізання спеціальним ферментом з алкілірованої основи групи – СН3 Пряма фотореактивація – фермент фотоліаза відновлює піримидінові димери. Темнова репарація – забезпечує відновлення УФ- пошкоджень ДНК. Ексцизіонна репарація – вирізання пошкоджених ділянок ДНК (АР-сайтів).
 реактивація Уейгла (індукція додаткових репаративних...")
Індукована репарація: а) реактивація Уейгла (індукція додаткових репаративних ресурсів клітин, які піддалися дії УФ- випромінювання), б) SOS – репарація (включення додаткових репаративних ресурсів при значному пошкодженні клітини). Репарація неспарених нуклеотидів - направлений рух комплекса репаративних білків по одному ланцюгу ДНК. Рекомбінантна репарація – відновлення структури ДНК, яке відбулося під впливом мутагенів (Н., - γ випромінювання). Відбувається при наявності непошкодженої гомологічної молекули ДНК.
 1. Види...")
Закон гомологічних рядів спадкових форм мінливості (М.І.Вавілов,1920) 1. Види і роди, які генетичне близькі, характеризуються подібними рядами спадкової мінливості з такою правильністю, що знаючи ряд форм у межах одного виду, можливе передбачити існування паралельних форм у інших видів та родів. Чим генетичне ближче організми в загальної системі родів та видів, тим повніше схожість в рядах їх мінливості. Свій закон М.І.Вавілов виразив формулою: G1 ( a + b + c ……….), G2 ( a + b + c ……….), G3 ( a + b + c ……….), де G – різні види (роди) організмів, a , b, c – різні вариабельні ознаки.
 організмів взагалі характеризуються певним циклом гомо...")
2. Цілі родини (групи) організмів взагалі характеризуються певним циклом гомологічних форм мінливості, які спостерігаються у всіх родів та видів, які відносяться до даної систематичної групи. Закон гомологічних рядів спадкових форм мінливості відображає загальну направленість мутаційного процесу у всіх живих організмів, яка визначається: а) універсальністю генетичного коду, загальною схемою організації генів, процесами реалізації генетичної інформації; б) хромосомною організацією спадкового матеріалу; в) однаковими для еукаріот процесами ділення клітин; г) гомологічними механізмами рекомбінування, мутування; д) гомологічними процесами утворення статевих клітин та запліднення…

Біологичні антимутагенні механізми На популяційному рівні: - зниження репродуктивності; На организменому рівні: - вренатальна смертність; передчасна постнатальна смертність. На клеточном уровне: Элиминация гамет и зигот; Парность хромосомного набора. На молекулярно-генетичному рівні: повтори генів; триплетність; репарація.

Схожі презентації











Категорії











