Презентація на тему:
Иммуноглобул_
Завантажити презентацію

. Будова і властивості. Гени імуноглобулінів. Біосинтез...")






, що зв’язаний N-глікозидни...")











 може набувати форми зірки, а...")












")





 універсальність: антитіла можуть...")

 генів імуноглоб...")
 генів імуноглобулінів 1) сайт-специфічна рекомб...")



 Стадії 0 A B C D E F")
 0-лімфоїдна стовбурова кровотворна клітина...")
...")





 генів V-об...")











 =(diversification) імуноглобулінів...")






Иммуноглобул_
Завантажити презентаціюПрезентація по слайдам:
. Будова і властивості. Гени імуноглобулінів. Біосинтез...")
Антитіла (antibody=AB). Будова і властивості. Гени імуноглобулінів. Біосинтез антитіл.

Тема 4. Антитіла будова і властивості.

Антитіла – продуковані В- лімфоцитами глікопротеїни, відкриті як γ- глобулінова фракція сироватки крові, що здатні специфічно взаємодіяти з антигеном. В 1972 році Родні Портер із Оксфорду та Джеральд Едельман із Нью -Йорку отримали Нобелівську премію за вивчення структури імуноглобулінів. Згідно з їхніми дослідженнями було створено модель молекули антитіла.

Структура імуноглобулінів Н-важкі поліпептидні ланцюги 5 класів ланцюгів- відповідні класи імуноглобулінів α-Ig A підкласи α 1, α 2 γ-Ig G підкласи γ1, γ2, γ3, γ4 δ-Ig D ε-Ig E μ-Ig M підкласи μ1, μ 2 L-легкі поліпептидні ланцюги 2 типи- κ, λ Підтипи 1κ і 4 λ

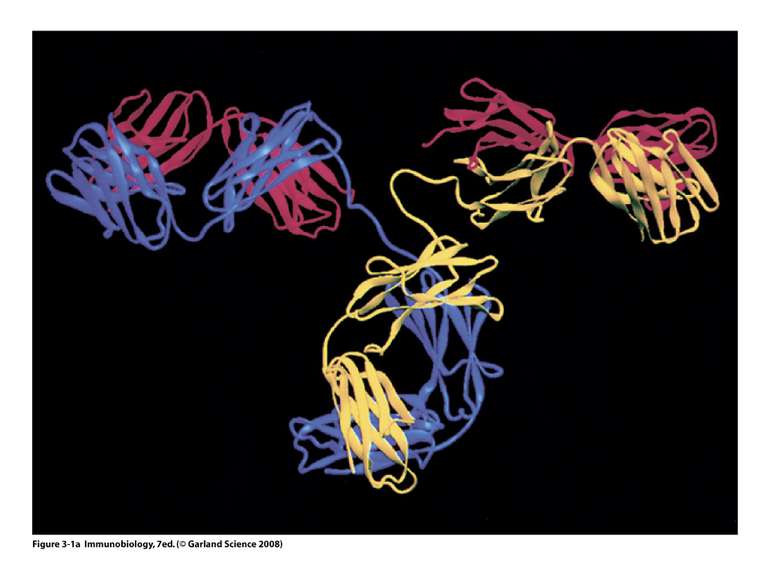
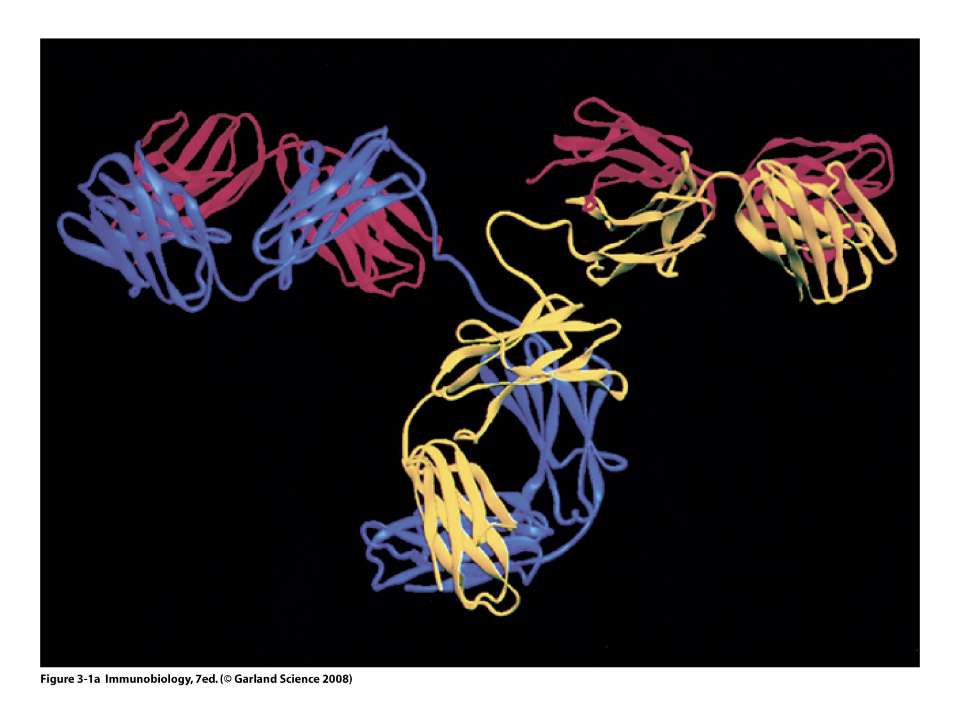
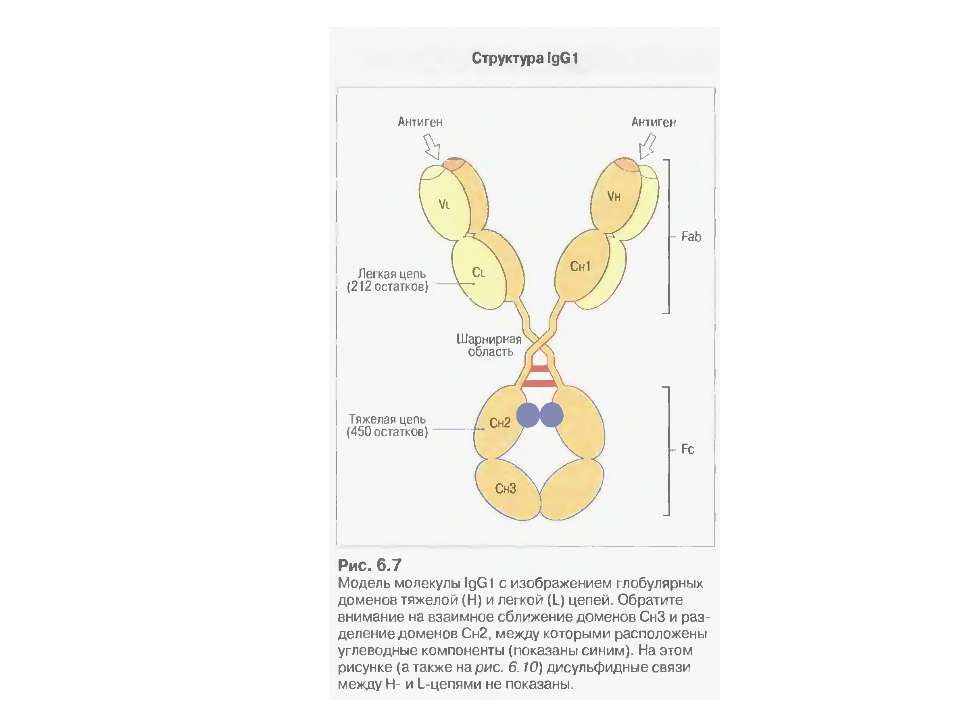
Будова антитіла


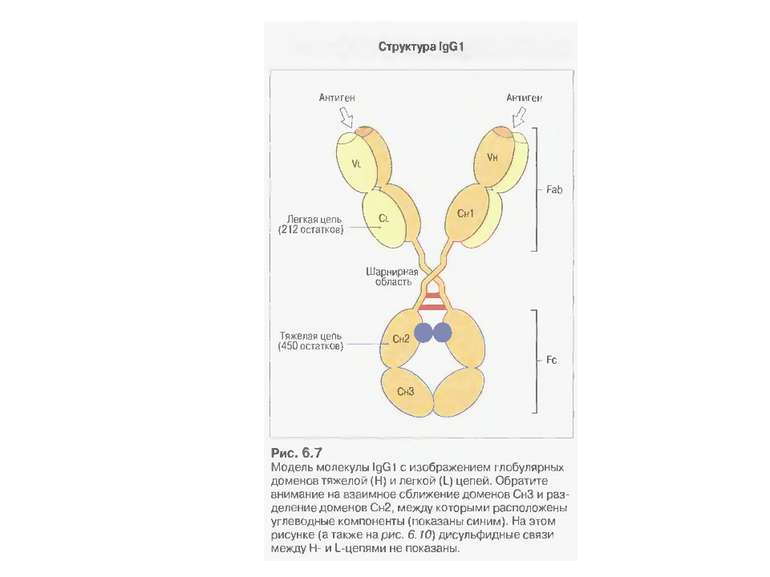
Будова антитіла Структура IgG


, що зв’язаний N-глікозидни...")
Олігоцукор (N-aцетилглюкозамін, манноза,галактоза), що зв’язаний N-глікозидним зв’язком із залишком аспарагіну у Fc фрагменті Ig G

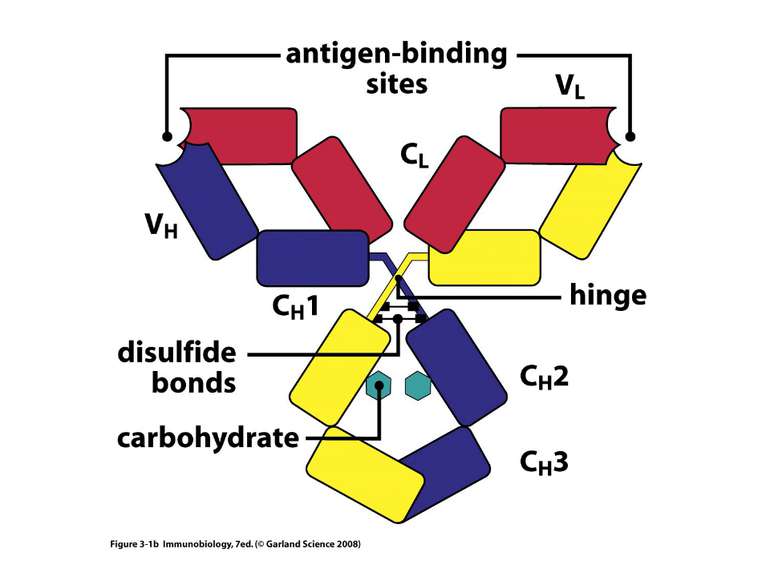
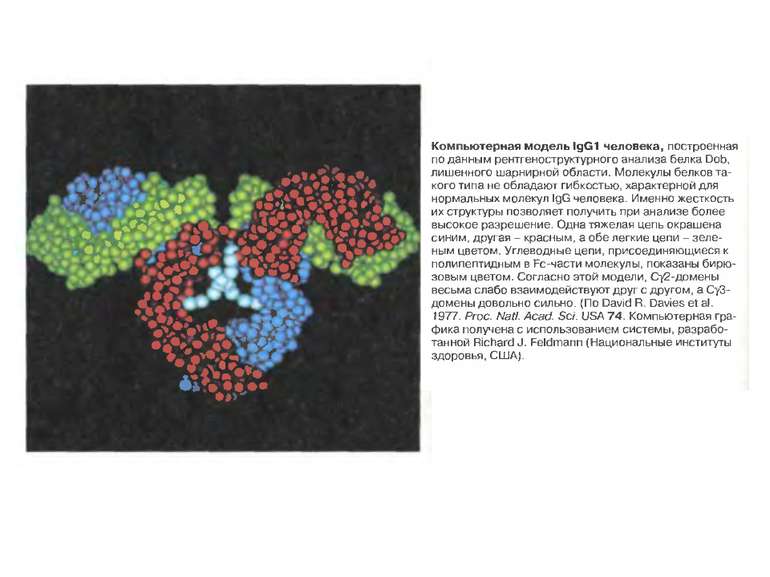
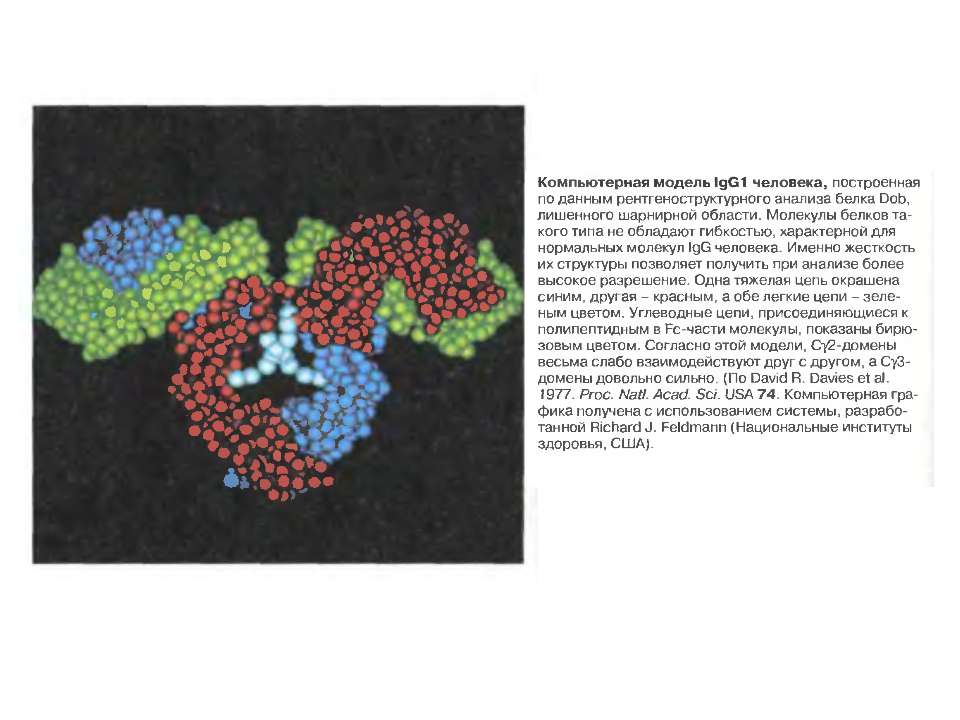
Надвторинна доменна організація імуноглобулінів

Надвторинна доменна організація імуноглобулінів

Надвторинна доменна організація імуноглобулінів (всередині-цілісна молекула білка, зліва-константні домени, справа – варіабельні домени)

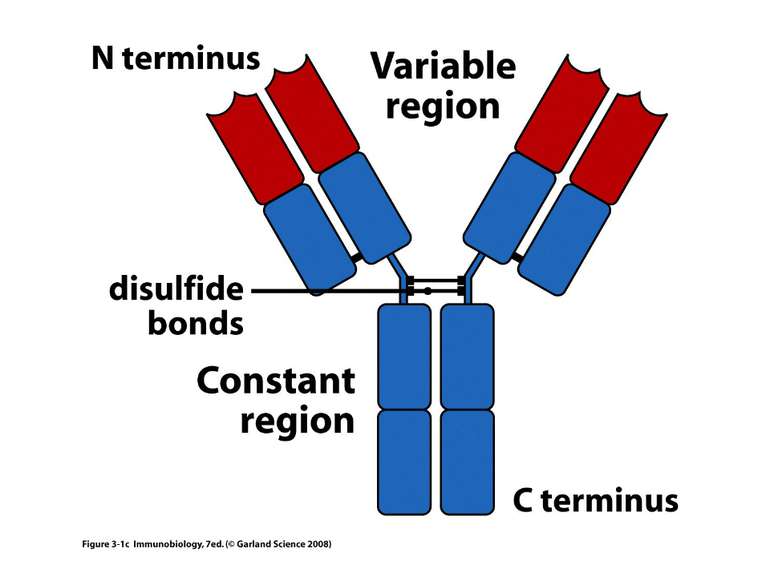
Типова молекула імуноглобуліну. Структурна організація легкого ланцюга імуноглобуліну

Типова молекула імуноглобуліну. Структурна організація легкого ланцюга імуноглобуліну

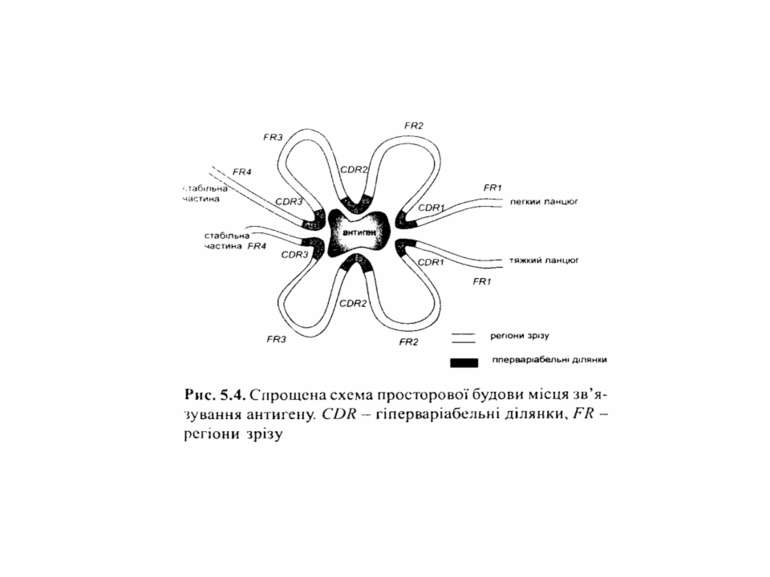
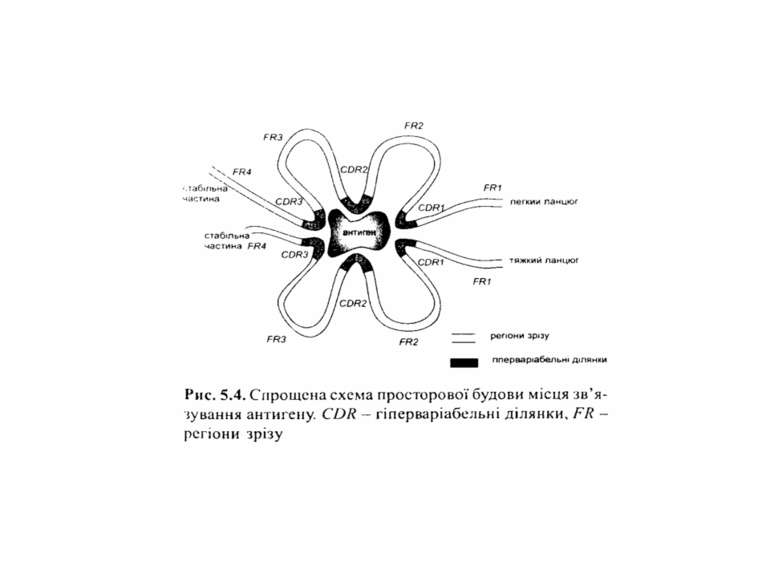
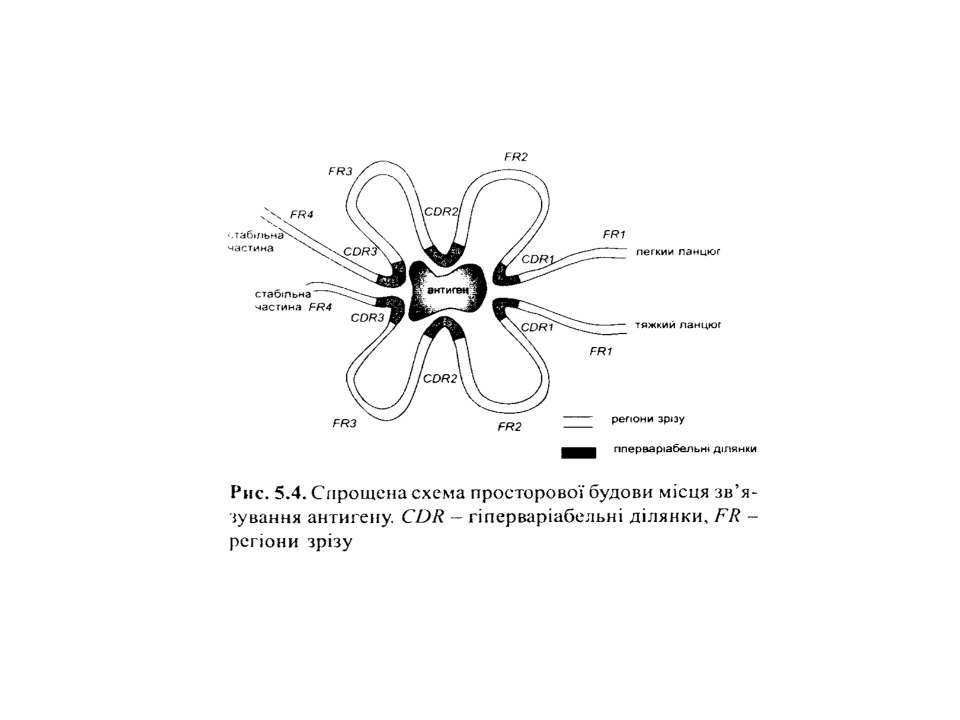
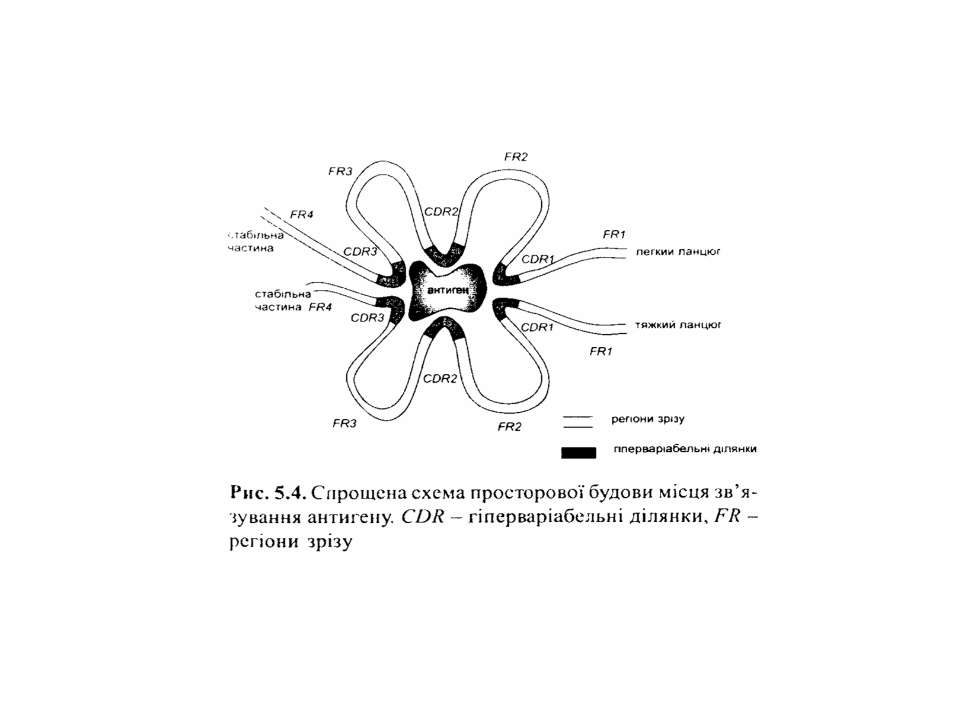
Будова імуноглобулінів CDR-complementarity determining regions

Будова імуноглобулінів CDR-complementarity - determining regions = HV- hypervariability regions; FR- framework region

Будова імуноглобулінів Cтруктура VL – варіабельного домену легкого ланцюга

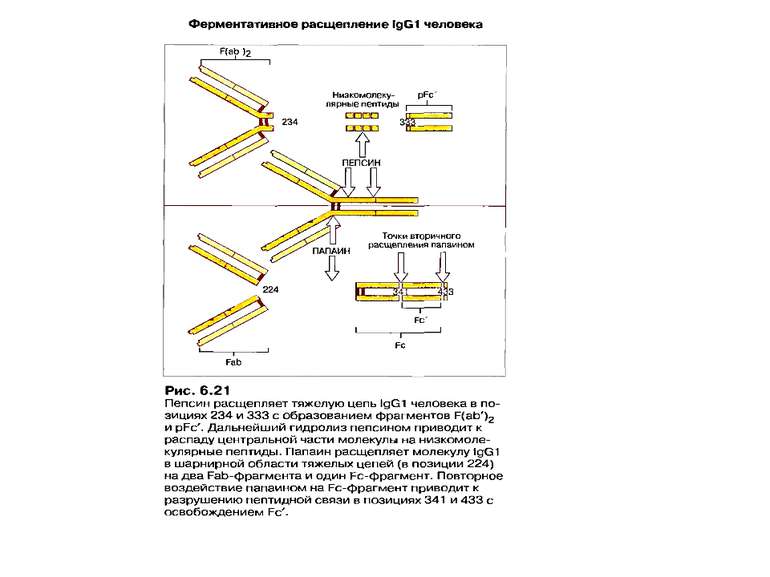
Протеолітичні фрагменти молекули антитіла

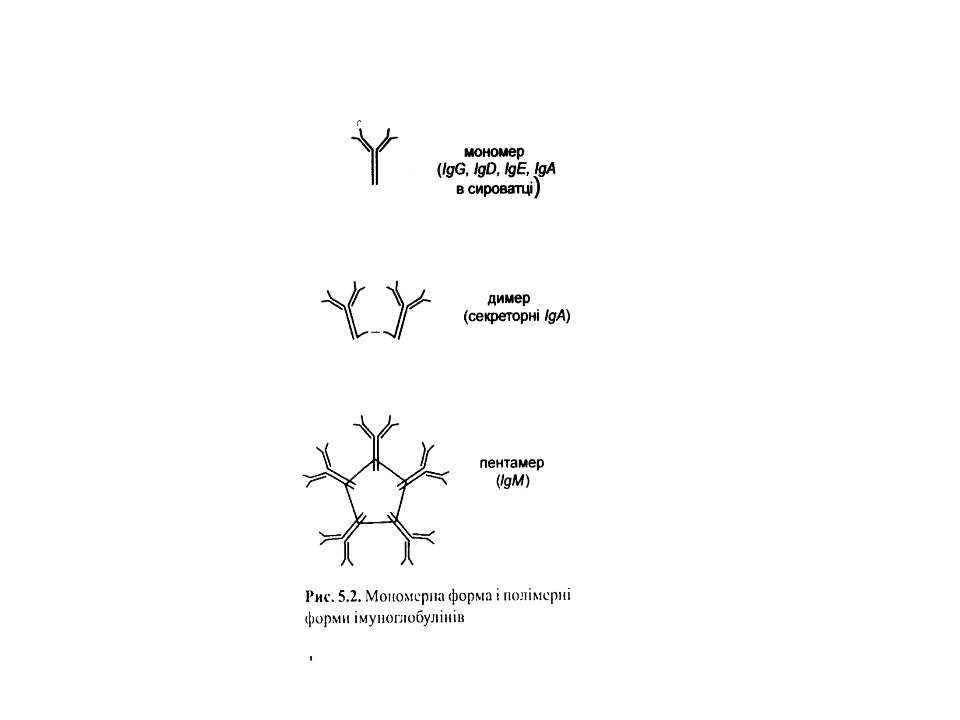

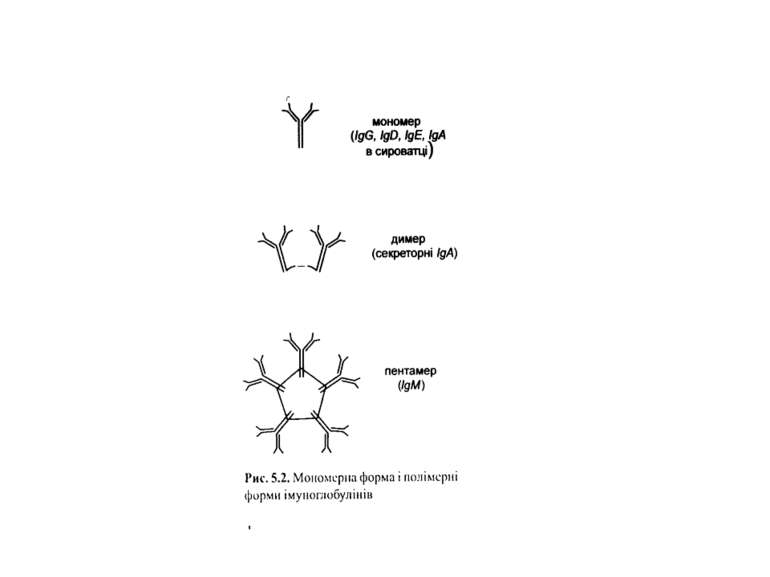
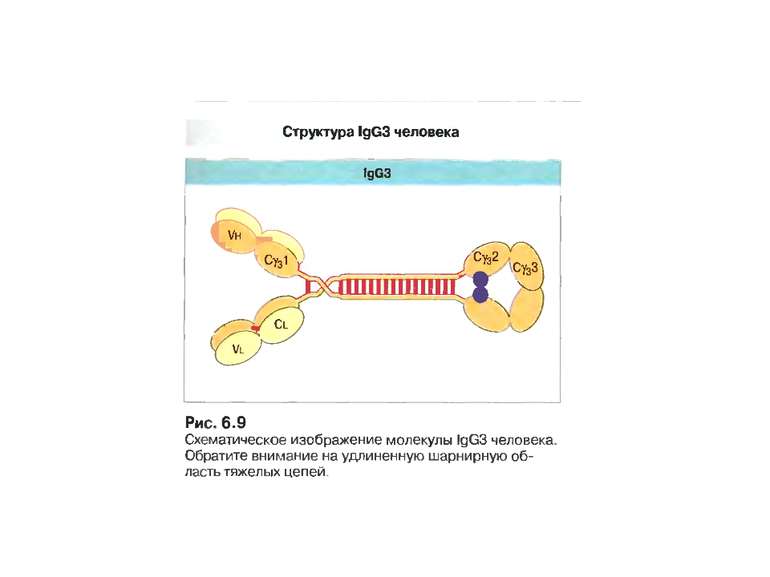
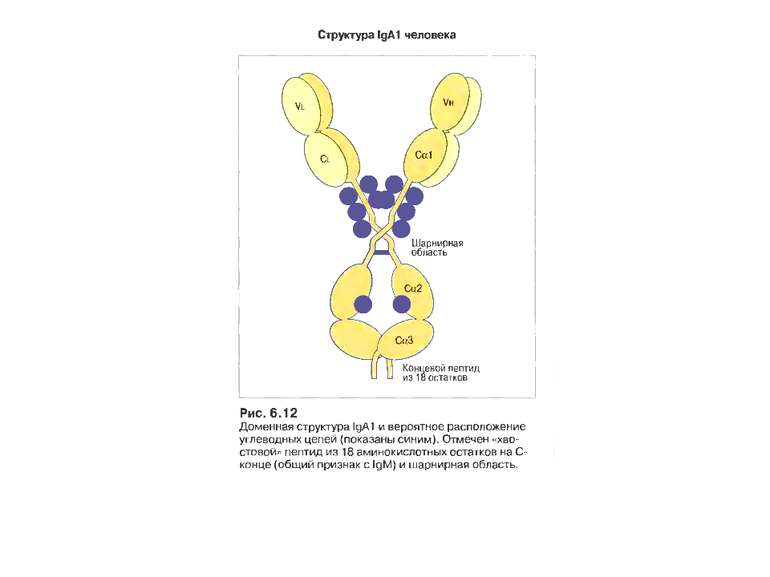
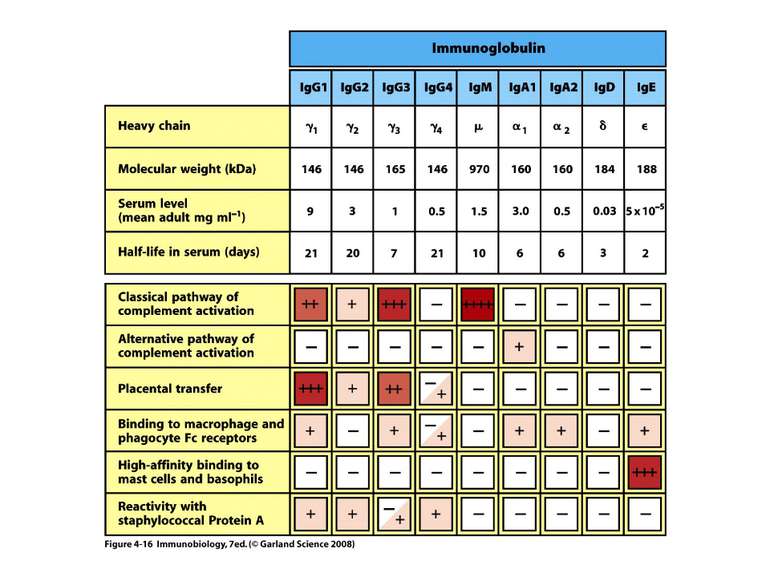
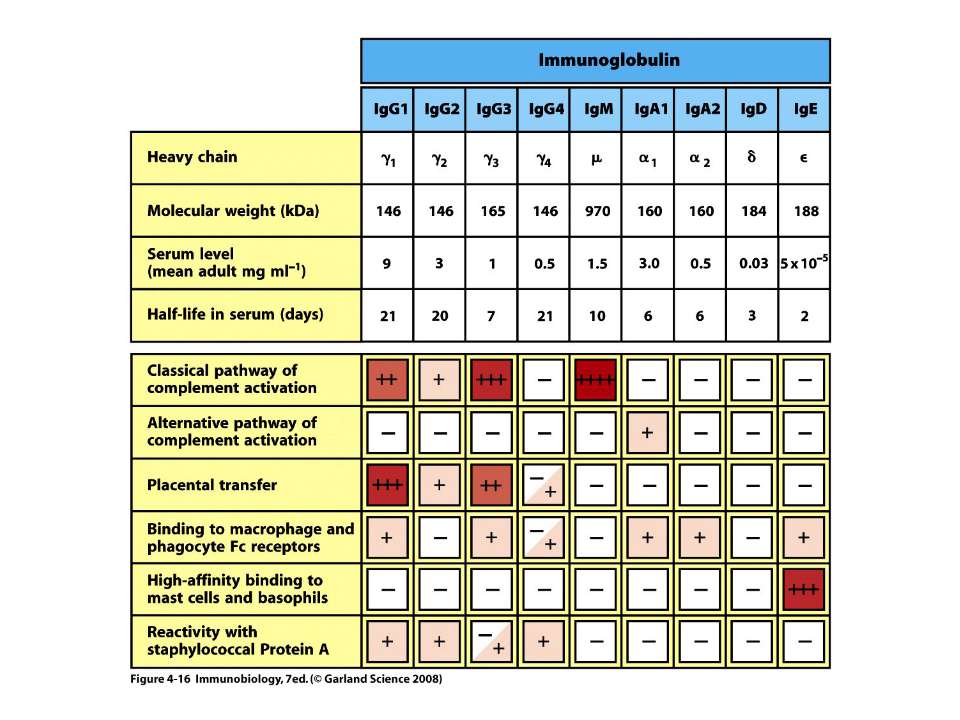
Класи імуноглобулінів

IgM 5-10% сироваткових Ig, Мм мономерної форми- 190 кДа, пентамерної форми - 970 кДа
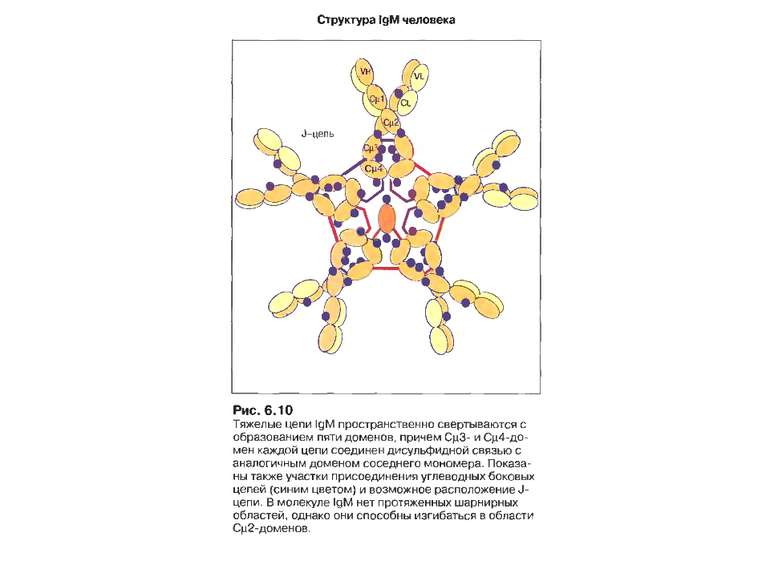
 може набувати форми зірки, а...")
Форми IgM Вільний IgM (не зв'язаний з антигеном) може набувати форми зірки, а IgM, зв'язаний з антигеном, що містить лінійно повторювані епітопи, набуває форми краба (на підставі мікрофотографій R. Feinstein і співавт.).
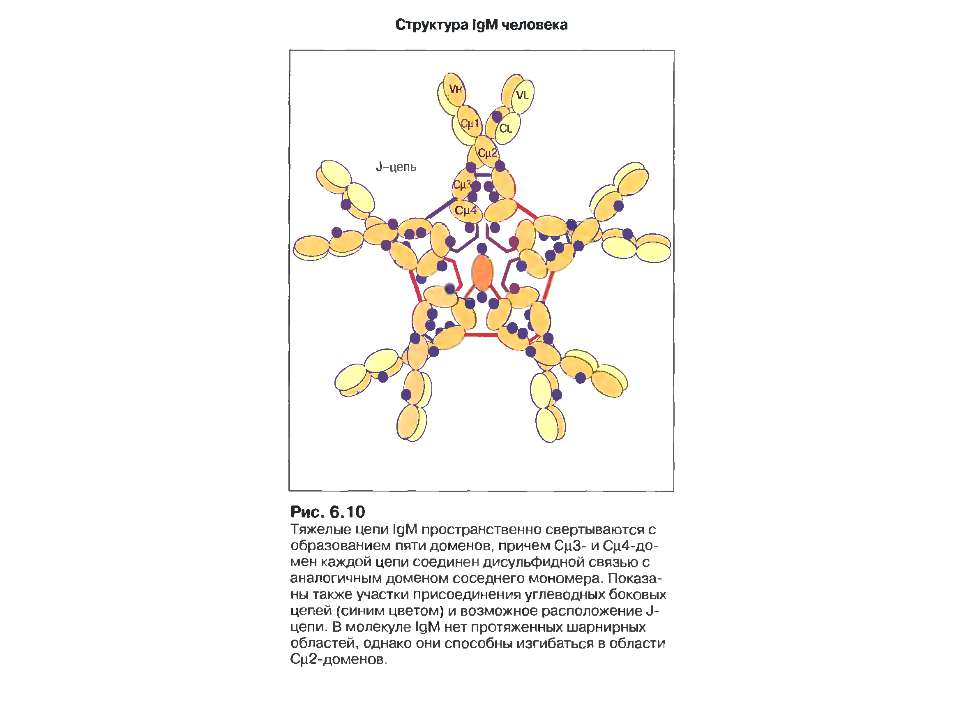

IgM

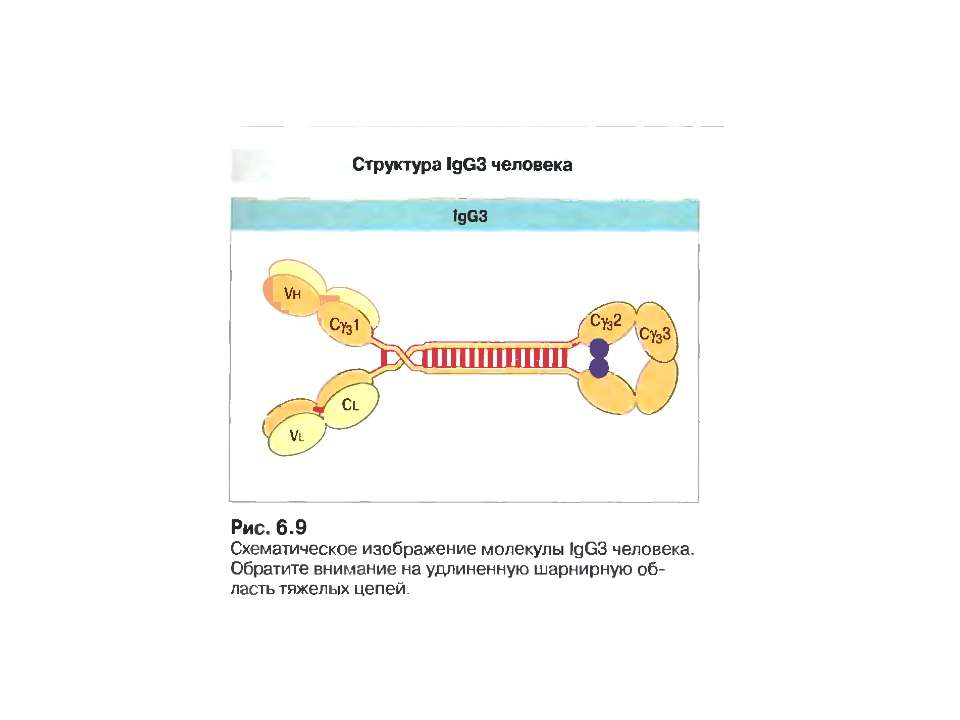
IgG 70-80% сироваткових Іg, Мм в середньому 150 кДа

Процес опсонізації

Процес опсонізації

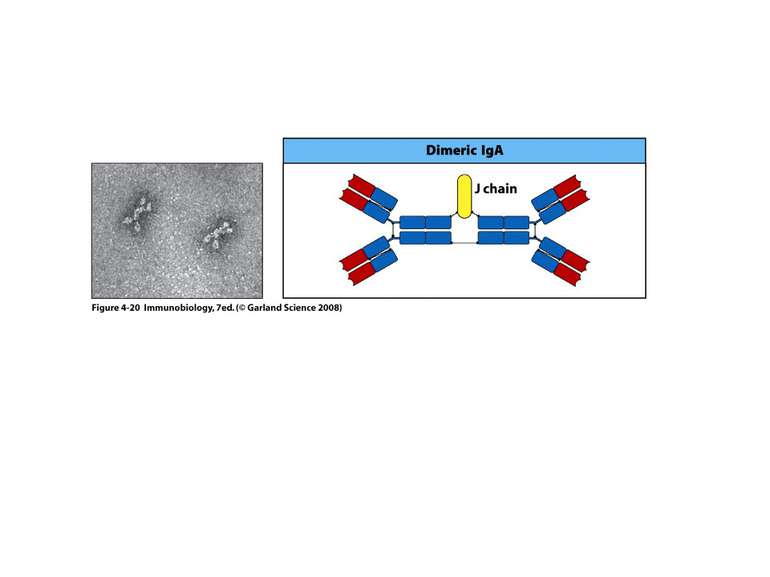
IgА 13-15 % сироваткових Іg, Мм в середньому 150 кДа
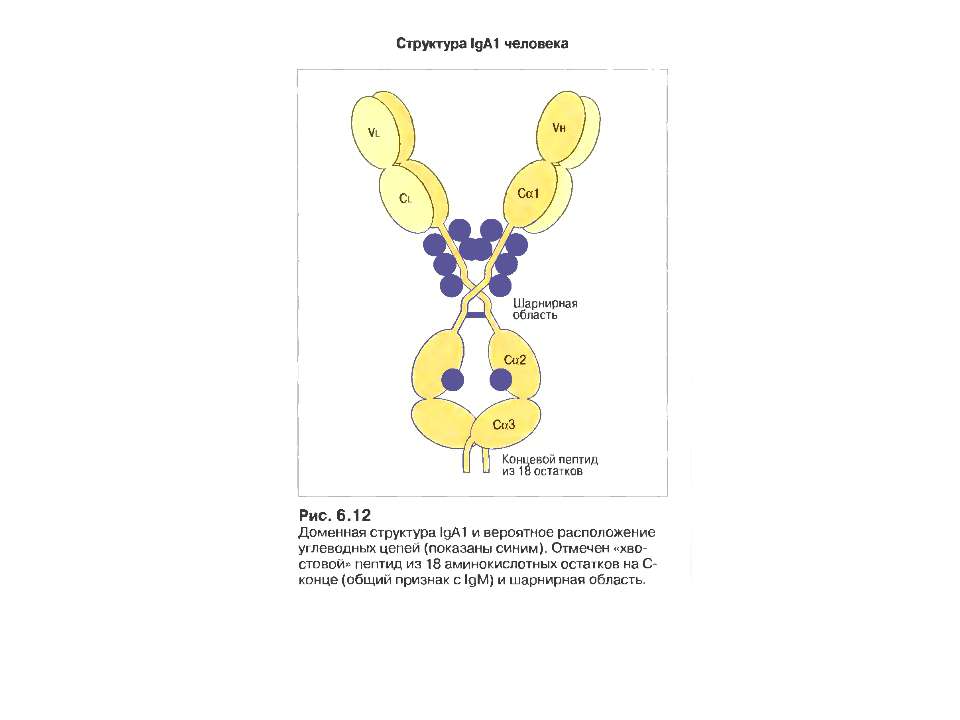
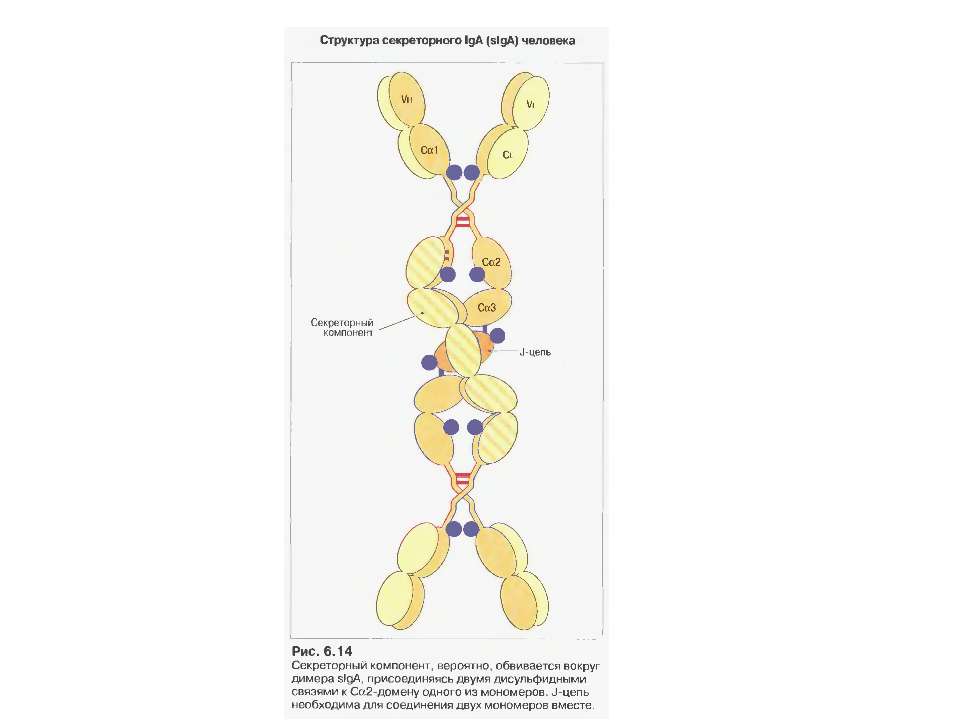
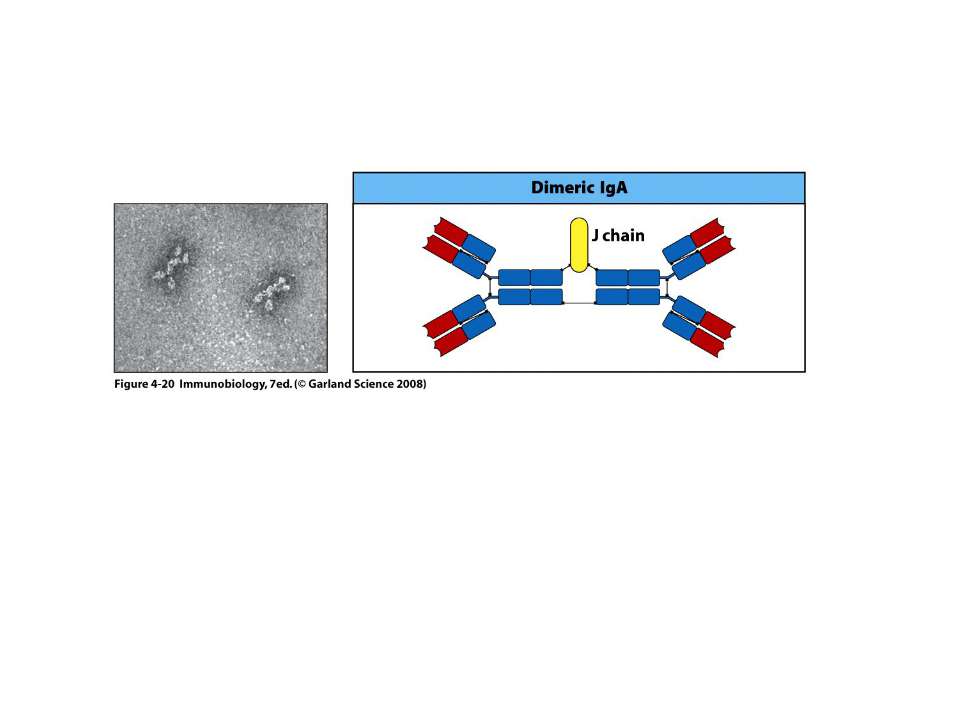

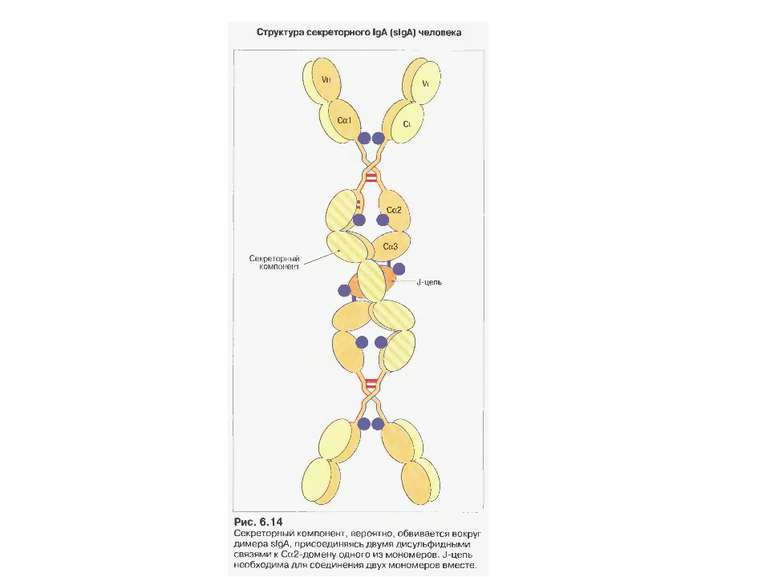
Механізм переносу sІgА через епітеліальну клітину

Механізм переносу sІgА через епітеліальну клітину

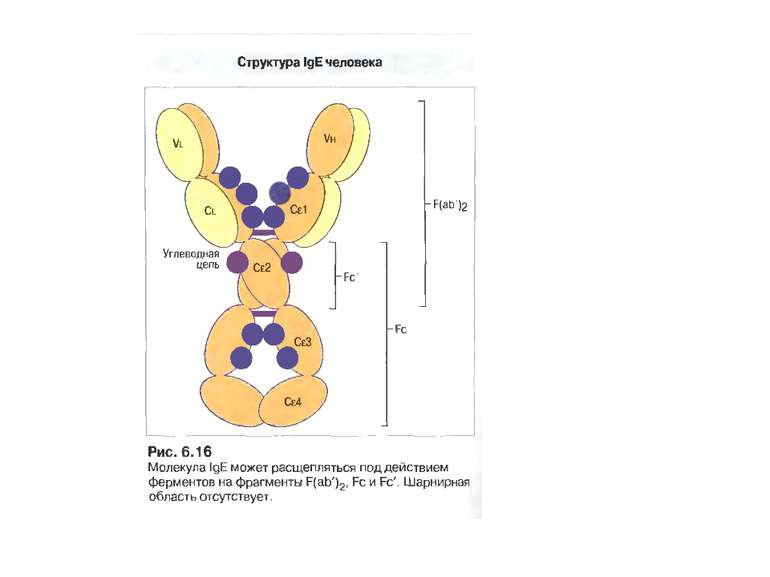
IgЕ 0,002 % сироваткових Іg, Мм в середньому 190 кДа
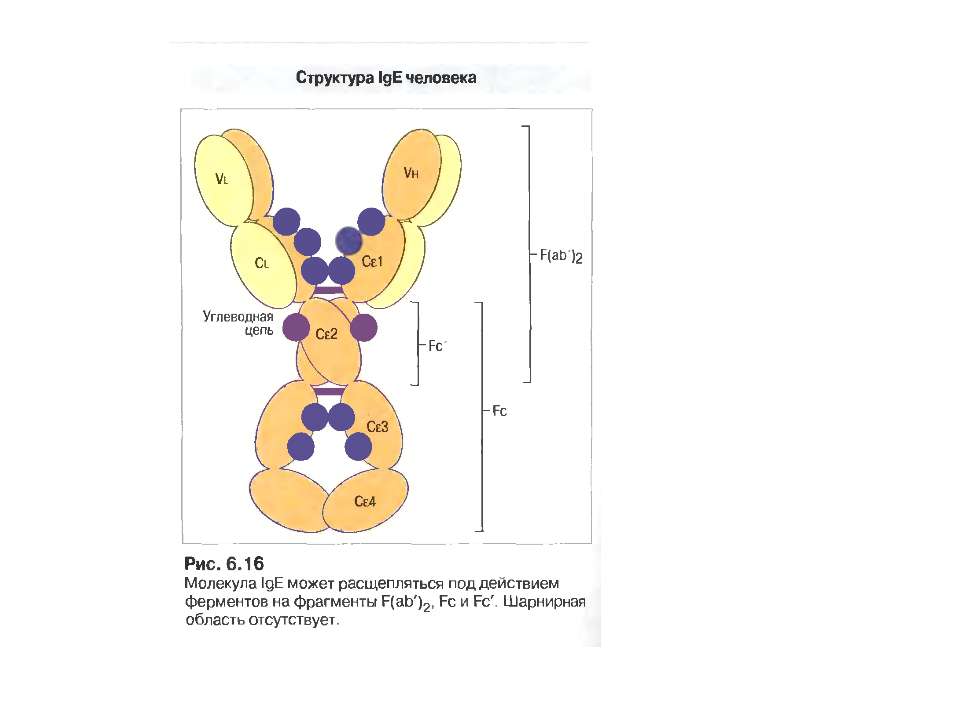

Функціонування IgE

Функціонування IgE

ІgD 2 % сироваткових Іg, Мм в середньому 185 кДа

Ig D
")
Середня концентрація у сироватці (мг/мл)

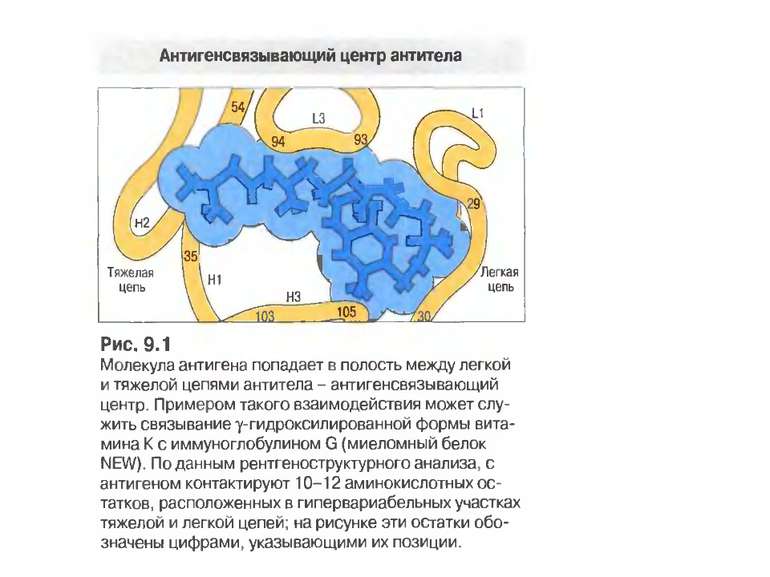
Будова активного центру антитіл 1. До складу активного центру антитіла входять гіперваріабільні амінокислотні залишки. 2. Зв'язок антигену з антитілом не ковалентний, а іонний, водневий , гідрофобний, Ван-дер-Ваальсівський. 3. Розміри активного центру - може розміститися 5-6 залишків амінокислот чи глюкози і розміри активного центру становлять, відповідно, приблизно 1,5х0,6х0,8 нм і можуть варіювати у різних антитіл. 4. Активний центр - це порожнина, заглиблена у Fv фрагмент приблизно на 1,5 нм. 5. У 70-ті роки 20 ст. було зроблено рентгеноструктурний аналіз мієломних імуноглобулінів, який підтвердив дані, отримані раніше, і показав, що активний центр антитіла - це порожнина, в утворенні якої беруть участь гіперваріабільні ділянки Fаb фрагменту, зближені у третинній структурі. За цими ж данними приблизно 10-12 амінокислотних залишків гіперваріабільних ділянок важкого і легкого ланцюга беруть участь у контактуванні з антигеном.

Будова активного центру антитіл


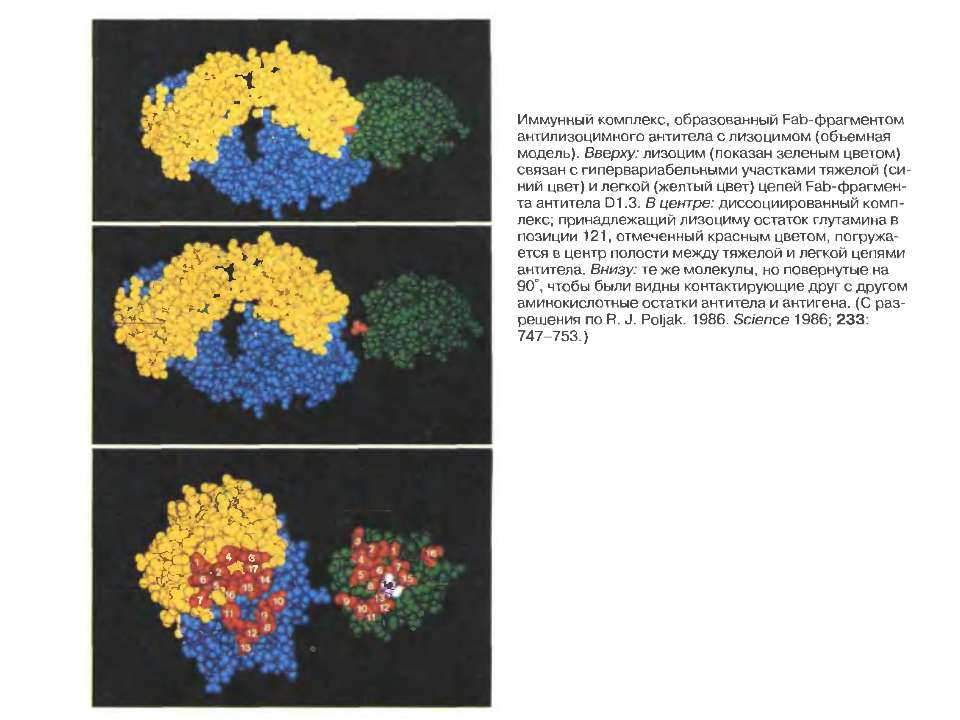

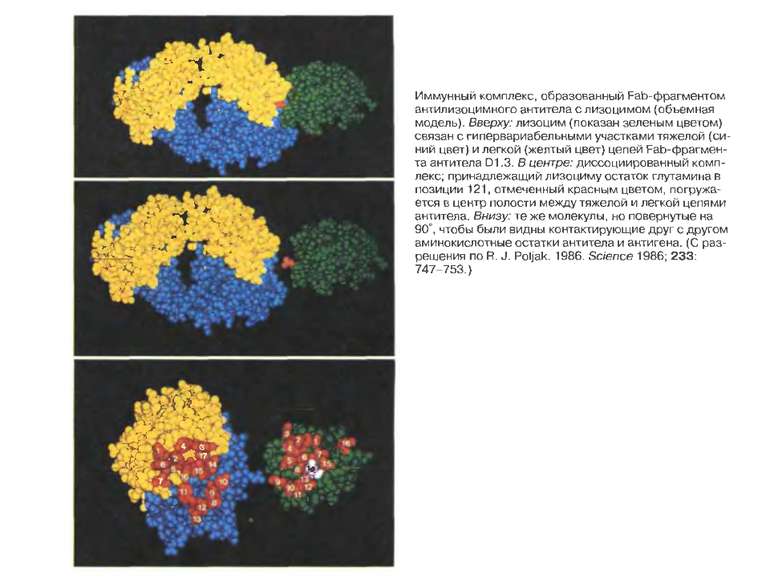
Будова активного центру антитіл Комплекс лізоциму курячого яйця з антитілом D1.3. Фіолетові-важкі ланцюги; легкі ланцюги-жовті; лізоцим-блакитний. Червоним позначено залишок глутаміну 121,що належить лізоциму , міститься між двома V- доменами і утворює водневі зв’язки, які стабілізують взаємодію АГ-АТ.

Будова активного центру антитіл
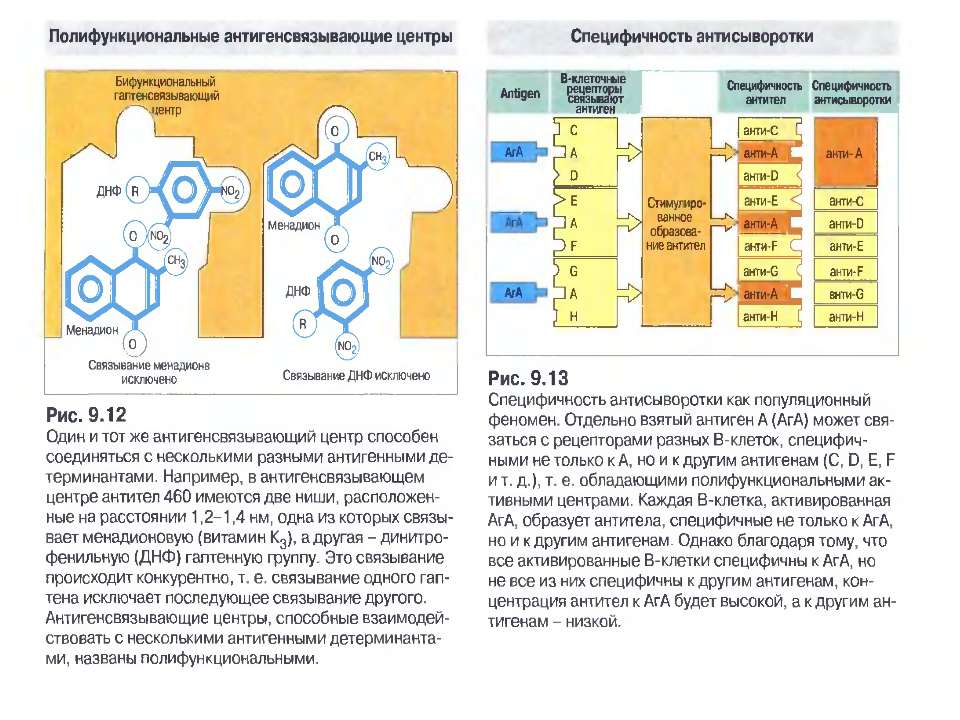
 універсальність: антитіла можуть...")
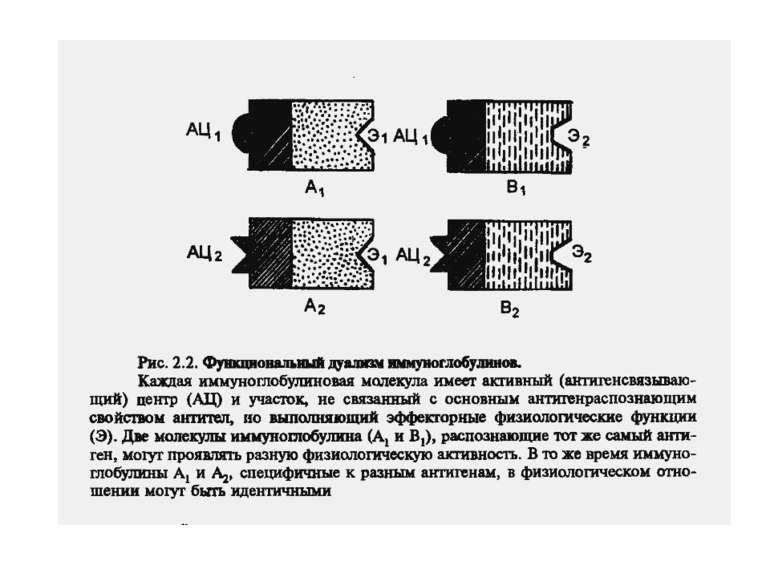
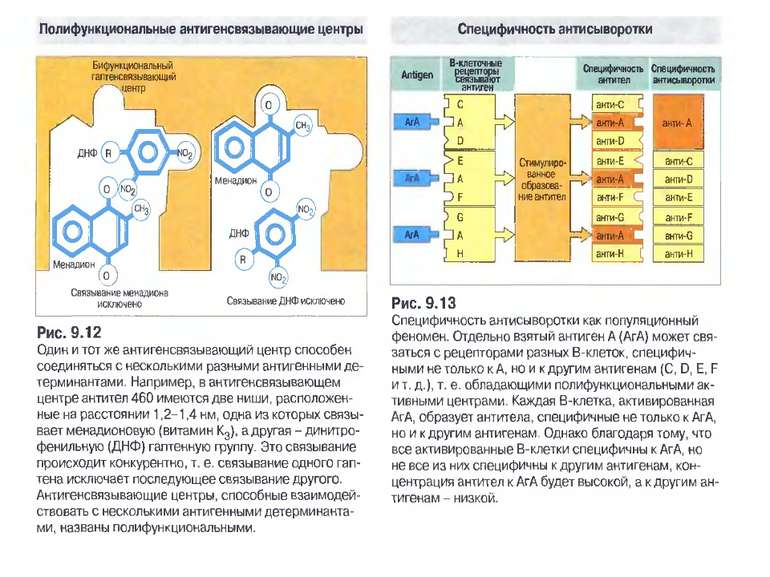
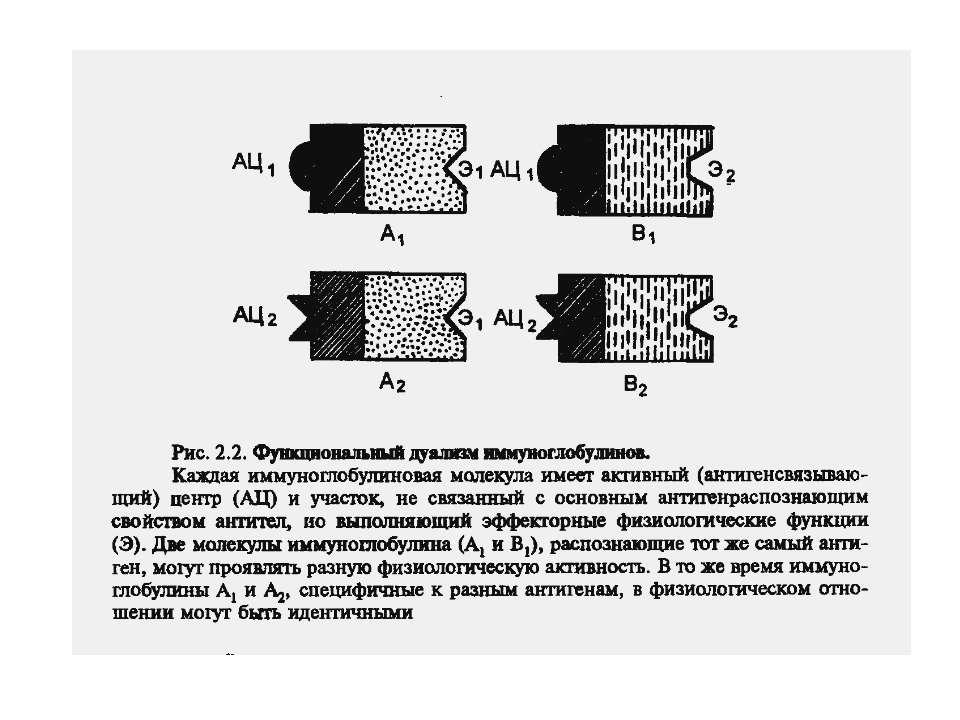
Чотири риси гуморальної імунної відповіді 1) універсальність: антитіла можуть бути виробленими проти будь-якого хімічного угрупування. Важко уявити, скільки повинно бути варіантів антитіл, якщо вважати, що кожний антиген потребує окремого антитіла; 2) специфічність: антитіла розрізняють о- чи p-положення функціональних груп, стереоізомери, білки, що відрізняються на один амінокислотний залишок; 3) гетерогенність: на один антиген виробляється багато типів (популяцій) антитіл, інколи більше 100 на одну антигенну детермінанту; 4) поліфункціональність: одне антитіло здатне зв'язувати більш, ніж один антиген. Поліфункціональність антитіл сприяє універсальності імунної відповіді, а гетерогенність - її високій специфічності.

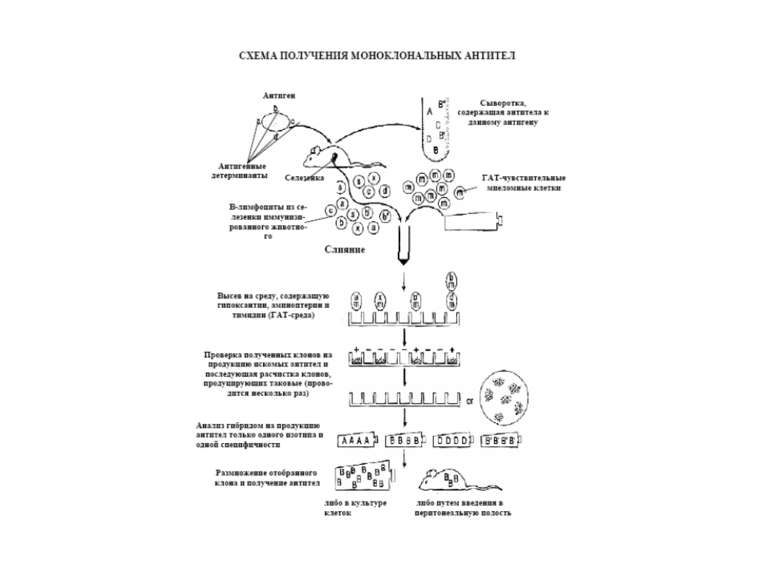
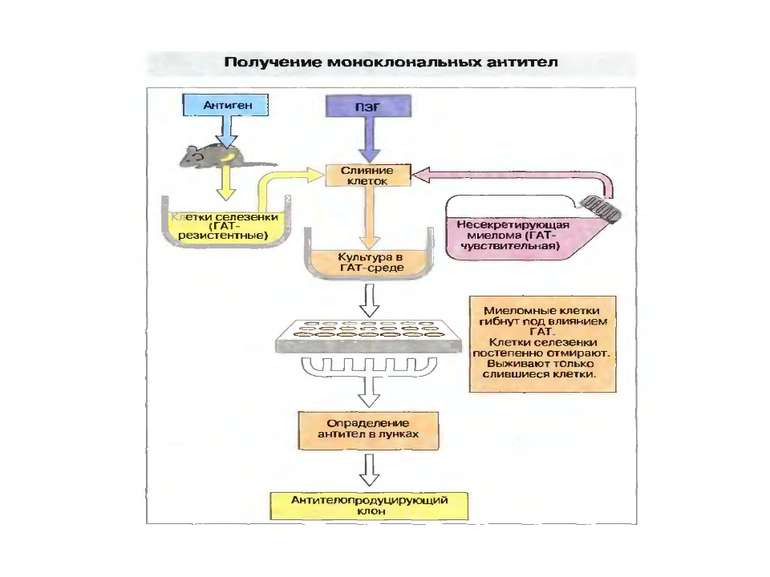
Тема 5. Гени імуноглобулінів. Біосинтез антитіл 1976 р. С. Тонегава і співавт.-довели можливість реаранжування генів 1987- Нобелівська премія за вивчення генів імуноглобулінів
 генів імуноглоб...")
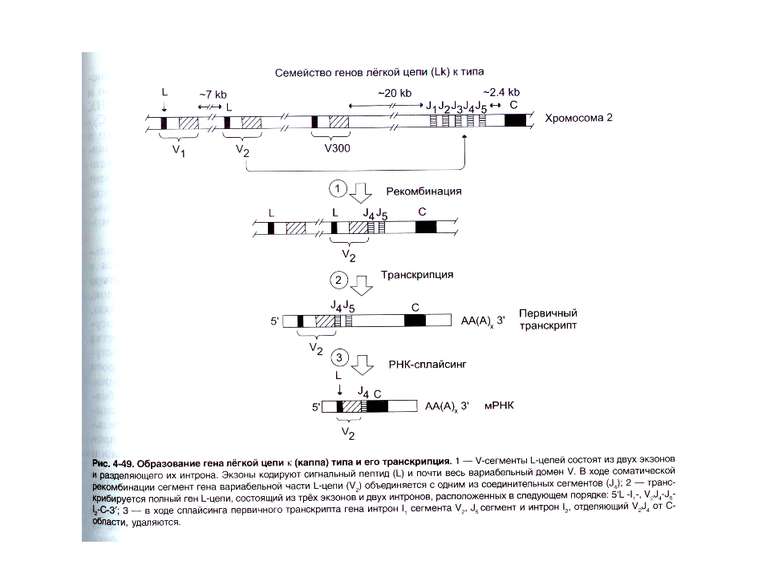
Гени імуноглобулінів. Біосинтез антитіл Три кластери (локуси) генів імуноглобулінів: Н, κ, λ. У людини – Н- 14 хромосома, κ- 2 хромосома, λ-22 хромосома. У миші- Н- 12 хромосома, κ- 6 хромосома, λ-16 хромосома. Входять- V, C гени важких і легких ланцюгів, J-гени (joining) , D-гени (diversity). Кластер генів λ -ланцюга . 30 V генів, 4 (6) пар J λ -і Сλ –генів-є зчепленими між собою. 2. Кластер генів κ –ланцюга. 40 V генів, 5 J-генів, 1 С κ -ген. Кластер генів Н-ланцюгів. Більше 1000 V- генів (функційні –51), 27 D-генів, 6 J-генів та С-гени всіх класів імуноглобулінів. V-сегменту передує ділянка ДНК, що кодує лідерний пептид (L).
 генів імуноглобулінів 1) сайт-специфічна рекомб...")
Реорганізація (реаранжування) генів імуноглобулінів 1) сайт-специфічна рекомбінація VJ ( для генів легких ланцюгів)-одна рекомбінаційна подія; 2) сайт-специфічна рекомбінація DJ (1) і V+DJ (2)- для генів важкого ланцюга- дві рекомбінаційні події; 3) транскрипція цілого блоку V(D) J- intron- С; 4) сплайсинг РНК з утворенням функціонального транскрипту V(D)JC; 5) трансляція; 6) посттрансляційне відщеплення лідерної послідовності.

Організація локусу генів зародкової лінії , що кодують імуноглобуліни людини

Транскрипційний промотор активується тільки після проходження рекомбінації, тобто транскрипція неперебудованої ДНК є неможливою

Схема перебудови генів імуноглобулінів: гени, що кодують варіабельні домени, утворюються в результаті сайт-специфічної рекомбінації
 Стадії 0 A B C D E F")
Стадії дозрівання В-лімфоцитів (1) Стадії 0 A B C D E F
 0-лімфоїдна стовбурова кровотворна клітина...")
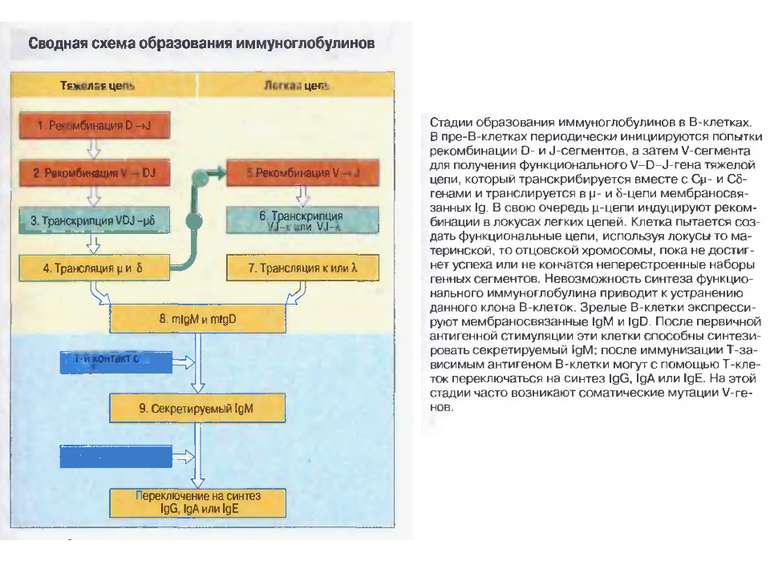
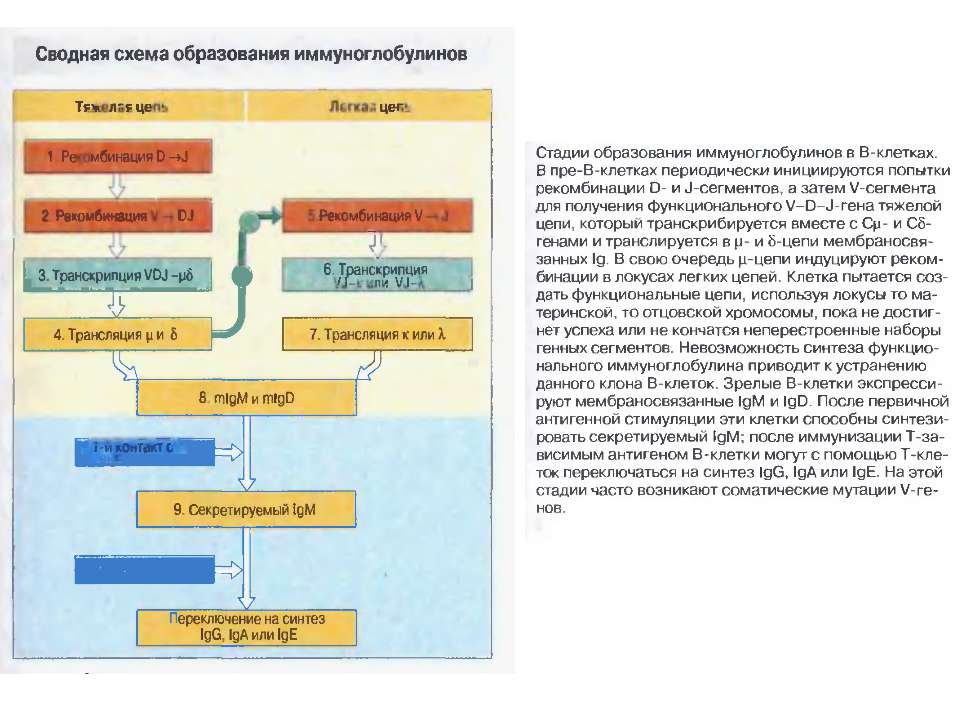
Стадії дозрівання В-лімфоцитів (2) 0-лімфоїдна стовбурова кровотворна клітина. А-ранні про-В-клітини-не мають рецепт. В клітин, початок реаранжування DJ-сегментів в локусі важкого (Н) ланцюга імуноглобулінів, може відбуватись в обох гомологічних хромосомах. В- пізні про-В-клітини- V- DJ реаранжування генів- підлягає алельному виключенню- тобто проходить лише в одній хромосомі. Отже на цій стадії перебудовуються гени і екс- пресується важкий ланцюг в комплексі з сурогатним легким, що є сигналом для позитивної селекції цих клітин. С- великі пре-В-клітини- експресують пре-В-клітинний рецептор, мають форму бластів, утворюють клони : 6 поділів, не працюють рекомбінази. D- малі пре-В-клітини, перебудова генів легкого κ–ланцюга, який експресується на мембрані разом з перебудованим μ- важким ланцюгом. Непродуктивність одного гена-включається інший, ні - перебудова генів в λ-локусі. E- незрілі В-клітини, мають сформовані Ig M-рецептори, але підлягають негативному відбору. На цій стадії В клітини виходять з кісткового мозку до селезінки і вступають на стадію дозрівання. F- зрілі наївні В-клітини, які рециркулюють (до зустрічі з антигеном) . Таким чином, перебудова генів та синтез білкового продукту пов’язана з: А) перебудовою важкого ланцюга Іg на стадії про-В-клітини Б) На стадії пре-В-клітин- перебудова легкого ланцюга Іg.
...")
Джерела різноманіття антитіл Виділяють 6 можливих джерел різноманіття Іg : 1) множинність гаметних генів (існування копій майже кожного генного сегмента); 2) соматичні рекомбінації (урізноманітнення за рахунок сайт- специфічної рекомбінації варіабельних ділянок молекули); 3) вбудовування (інсерції) та делеції додаткових нуклеотидів=junction diversity=урізноманітнення з'єднань; 4) механізм комбінаторної диверсифікації (утворення можливих варіантів імуноглобулінів); 5) соматичний мутагенез; 6) генна конверсія (активація роботи псевдо-генів).

Джерела різноманіття антитіл

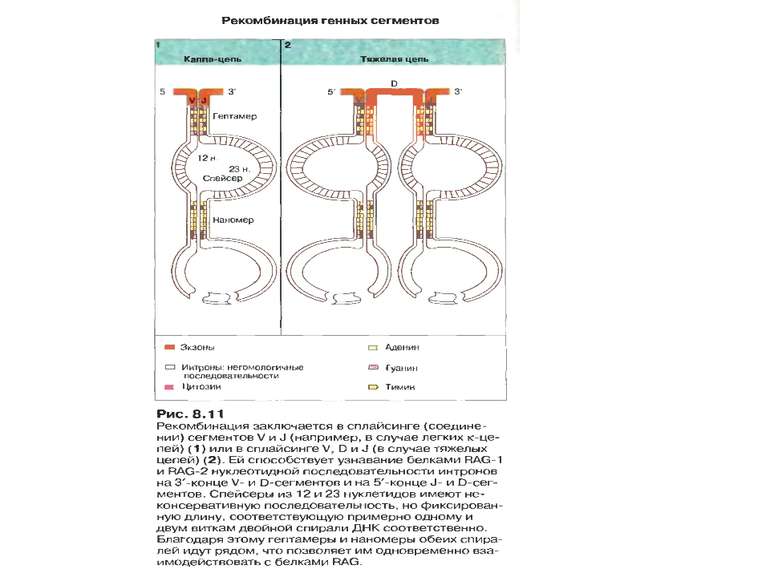
Механізми сайт- специфічної рекомбінації. Організація сигнальних послідовностей RSSs ( recombination signal sequences ) в процесі рекомбінації генів імуноглобулінів

Рекомбінація генних сегментів

Механізми сайт- специфічної рекомбінації. На етапах рекомбінації залучені: Продукти генів RAG1 і RAG2 -рекомбінази. Саме початок експресії цих генів є сигналом для початку перебудови генів імуноглобулінів. Крім того у складному процесі рекомбінації на етапі з'єднання кінців ДНК беруть участь білки, які працюють в системі репарації дволанцюгових розривів ДНК- [ консервативний процес NHEJ = nonhomologous DNA end-joining = негомологічне з'єднання кінців ДНК], модифікації кінців розривів і т. д., а саме: ДНК-залежна протеїнкіназа (DNA-PK) , білок Ки (гетеродимер Ки 70:80), який асоціює з DNA-PK, ендонуклеаза Artemis, процес лігування відбувається за участі ДНК-лігази IV та ДНК-репараційного білка XRCC4 , ендо- та екзонуклеази, термінальні дезоксинуклеотидил-трансферази (TdT), полімерази та інші.

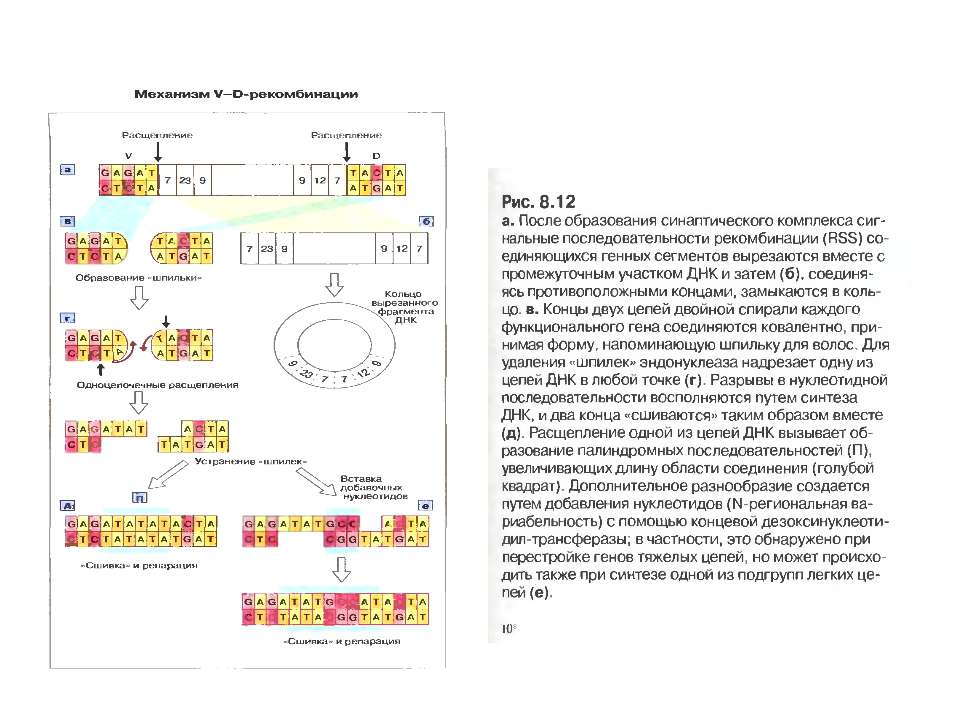

Механізм сайт- специфічної рекомбінації
 генів V-об...")
Механізм сайт- специфічної рекомбінації (RAG-залежна рекомбінація) генів V-області імуноглобулінів

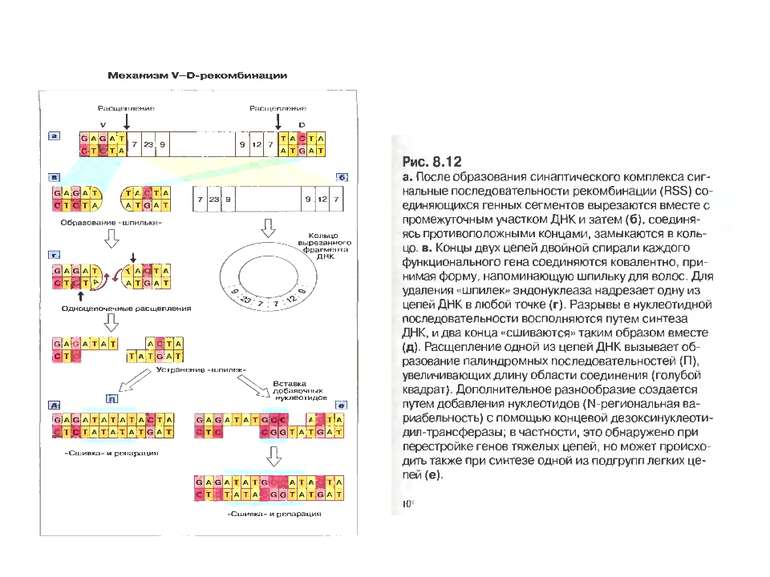
Схема приєднання нуклеотидів Р до рекомбінуючих генів D і J . Спочатку відбувається перерізання подвійної ДНК на межі сигнальних і кодуючих послідовностей з утворенням характерних "шпильок для волосся". Потім ці "шпильки" відкриваються в місцях, позначених ламаними стрілками. Виникають одноланцюгові відрізки ДНК, в яких два останні нуклеотиди комплементарні до нуклеотидів початкової послідовності (нуклеотиди Р-паліндромні). Тепер може відбутись і видалення, а також приєднання поодиноких нуклеотидів (не позначене на рисунку). Після приєднання комплементарних нуклеотидів відбувається відновлення кодуючої послідовності ДНК.

P-i N- нуклеотидні вставки при з’єднанні кодуючих сегментів генів імуноглобулінів

Будова імуноглобулінів CDR-complementarity determining regions

Легкі κ-ланцюги імуноглобулінів

Легкі λ-ланцюги імуноглобулінів
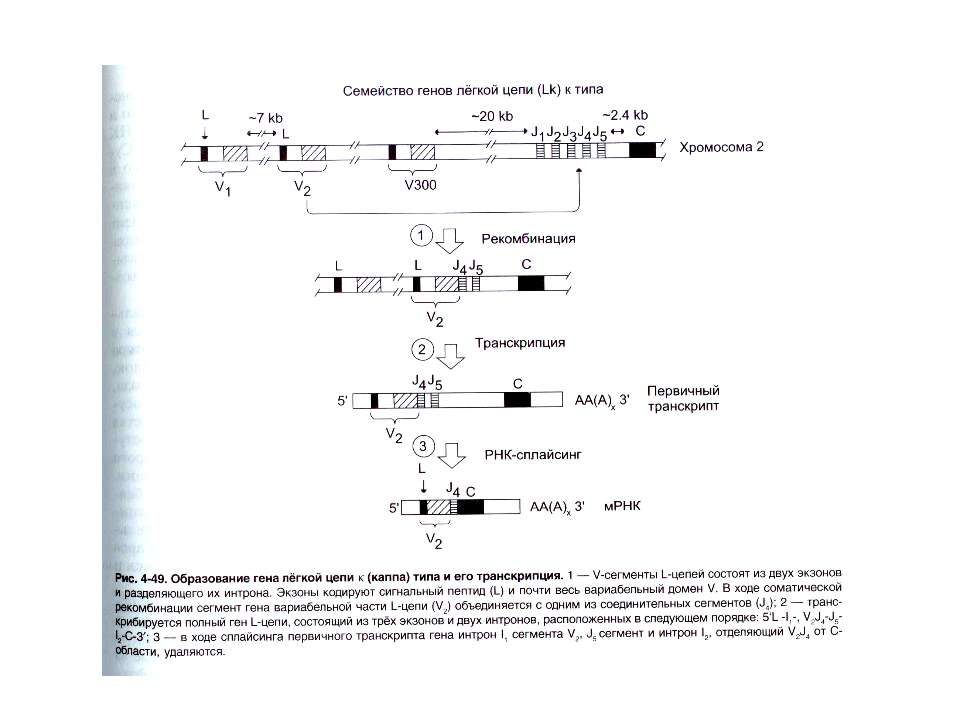
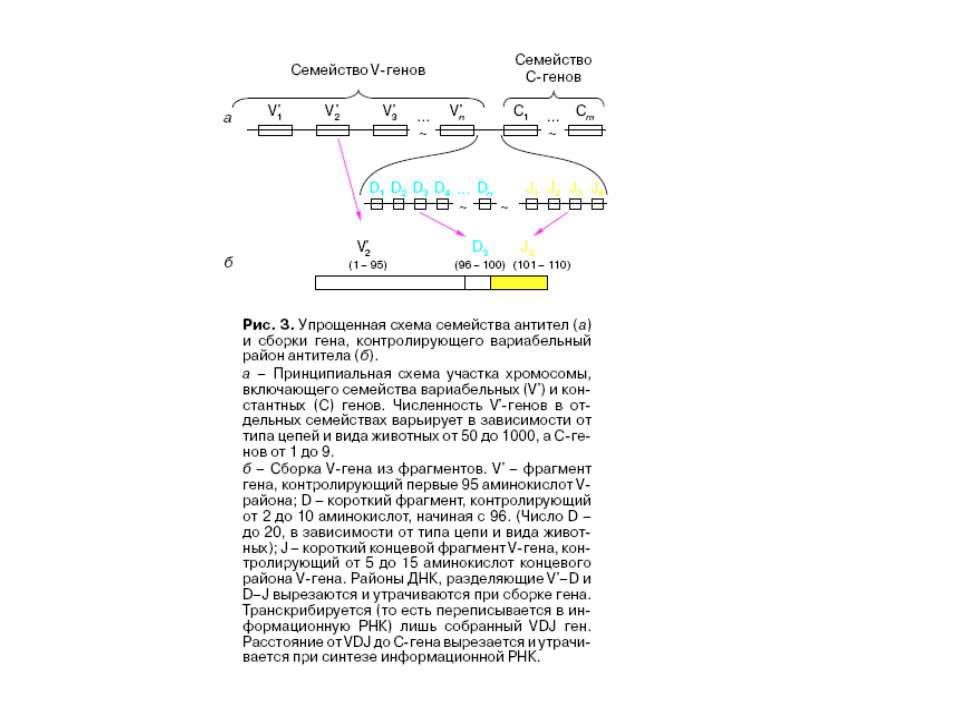

Схема збірки активного гена важкого ланцюга імуноглобулінів

Схема збірки активного гена важкого ланцюга імуноглобулінів

Важкі ланцюги імуноглобулінів μ δ γ3 γ1 γ2b γ2a ε α

Ізотипи імуноглобулінінів кодуються кластерами генів константних доменів важких ланцюгів

Схема збірки активного гена важкого ланцюга імуноглобулінів
 =(diversification) імуноглобулінів...")
Механізми диверсифікації (різноманітності) =(diversification) імуноглобулінів Комбінаторна диверсифікація (див. табл.кластерів генів) 1) L -ланцюги : λ -ланцюг - VJ: 30Vх 4J λ = 120; κ –ланцюг- VJ: 40V х 5Jк =200; 120 + 200= 320 варіантів L-ланцюга 2) Н ланцюги: DJ: 27D х 6J = 162; V(DJ): 51 х 162 = 8262; D ген має три вірогідні рамки зчитування: 8262 х 3 = 24786 варіантів Н-ланцюга. 3) Комбінація Н х L: 320 х 24786 = 7,9 х 106 4. Мутації у сайтах рекомбінації, генна конверсія, випадковість сайтів розщеплення за участі ендонуклеази Artemis, додавання Р і N-нуклеотидів - додають різноманітності >108

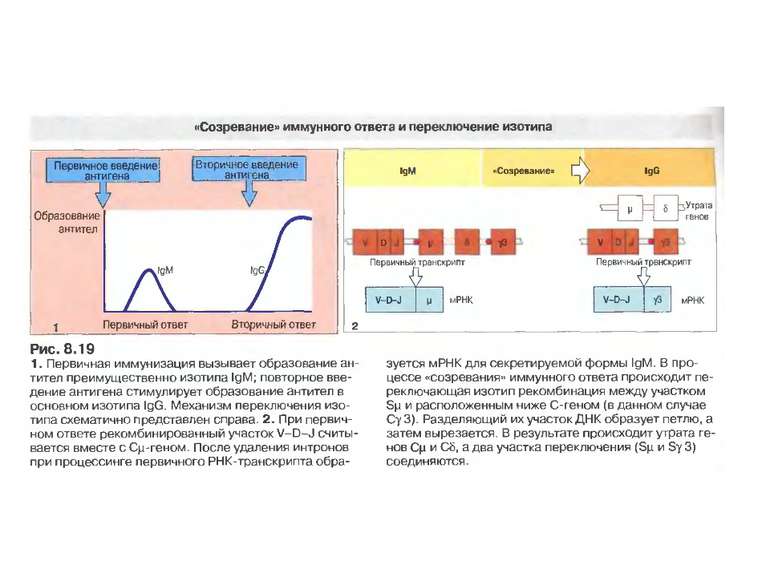
Соматичний гіпермутагенез

Алельне виключення

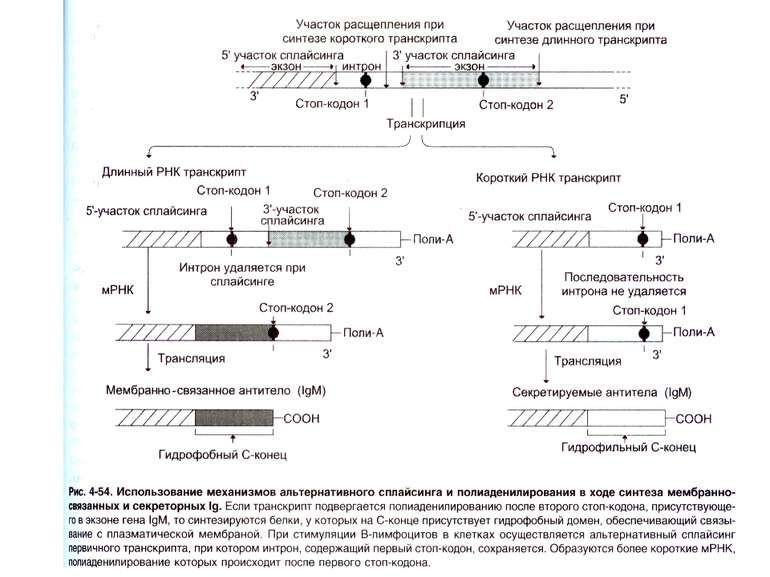
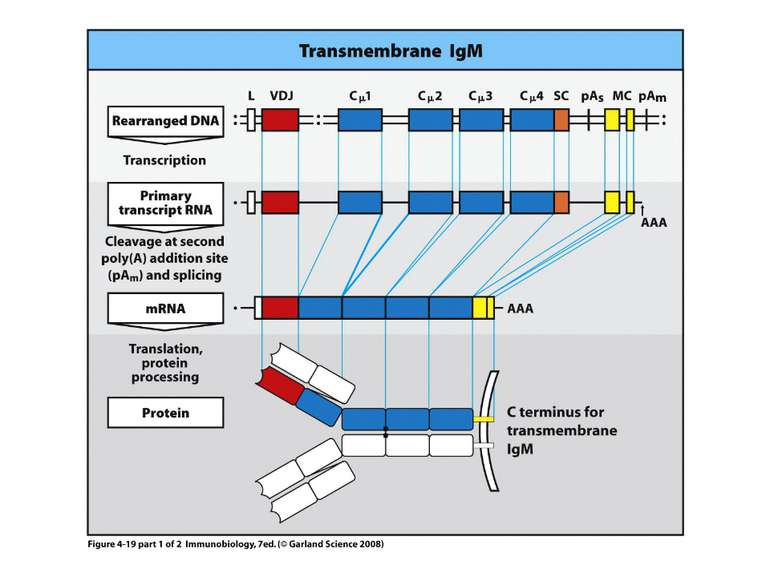
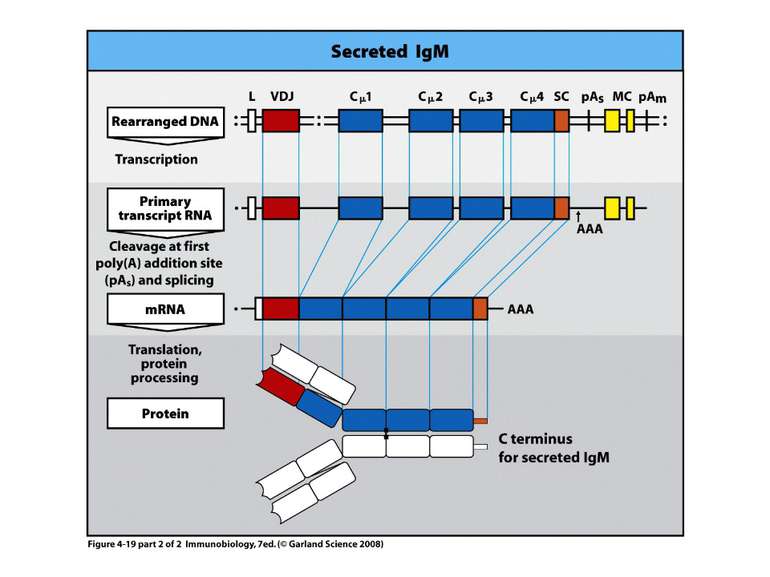
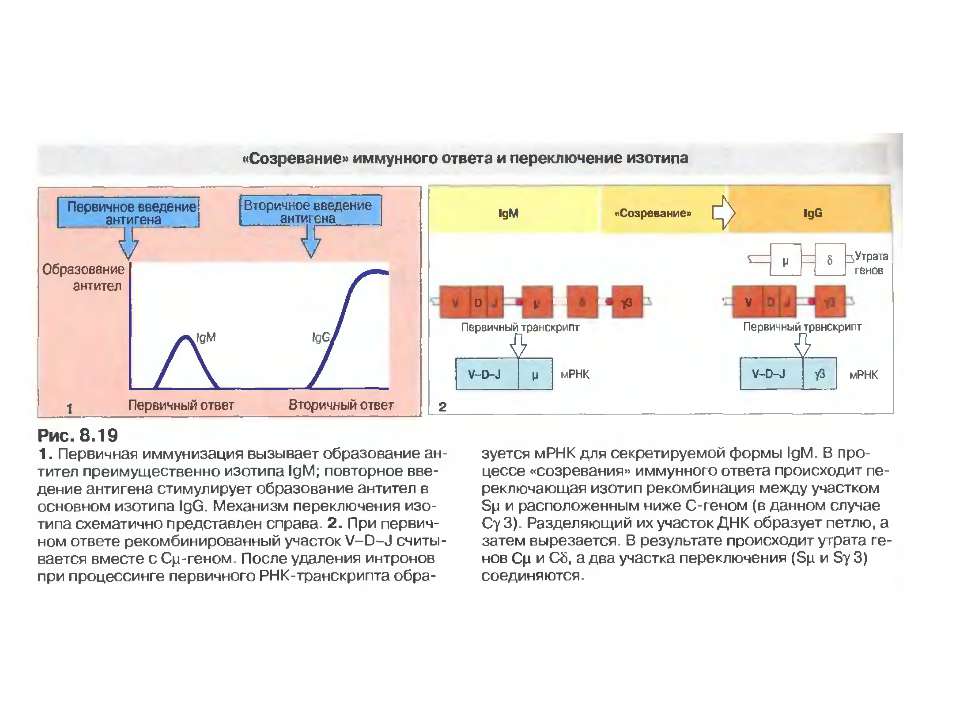
Переключення з мембранної на секретовану форму- на прикладі ІgМ

Переключення з мембранної на секретовану форму- на прикладі ІgМ

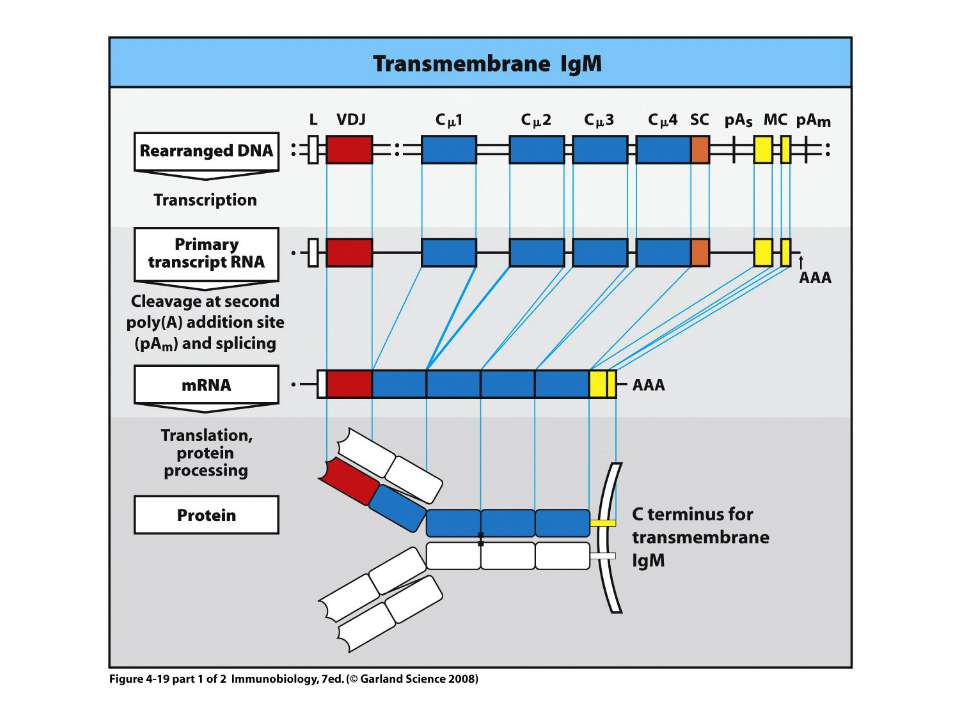


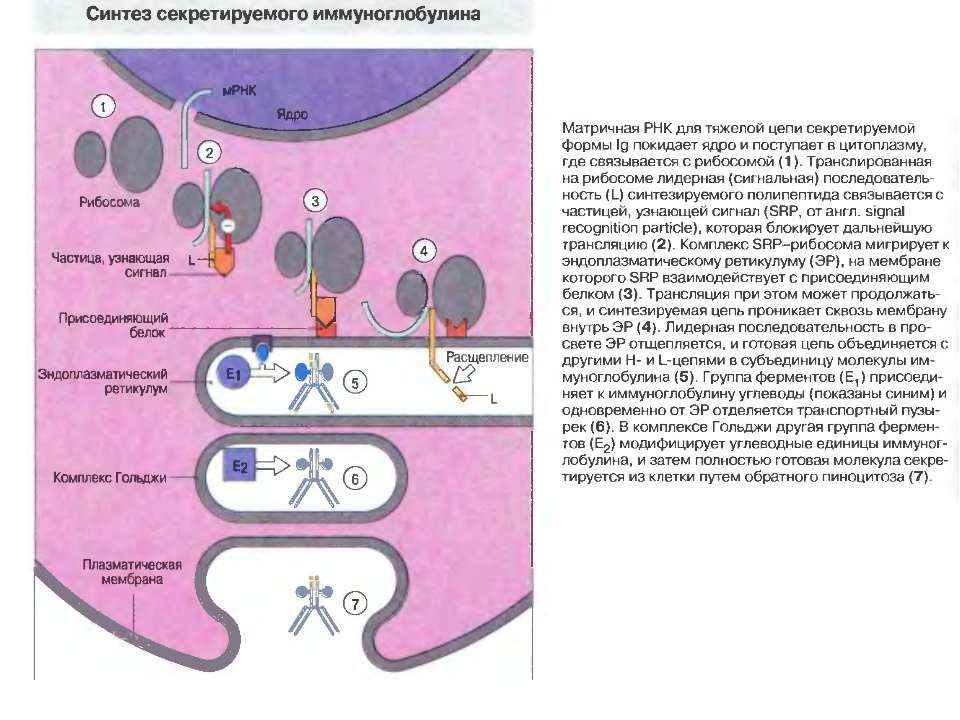
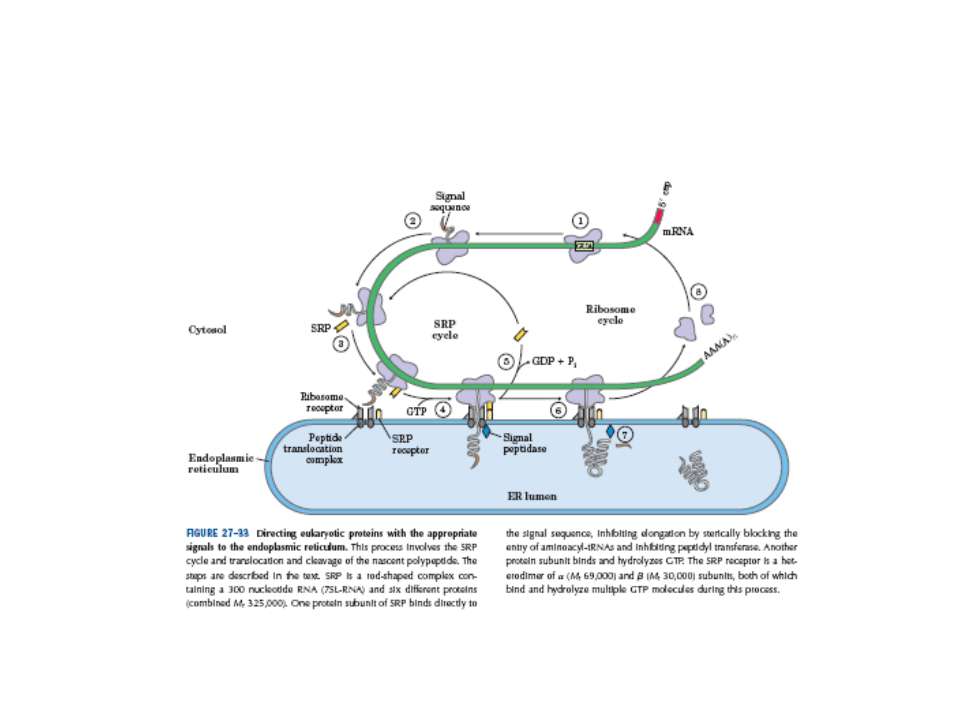

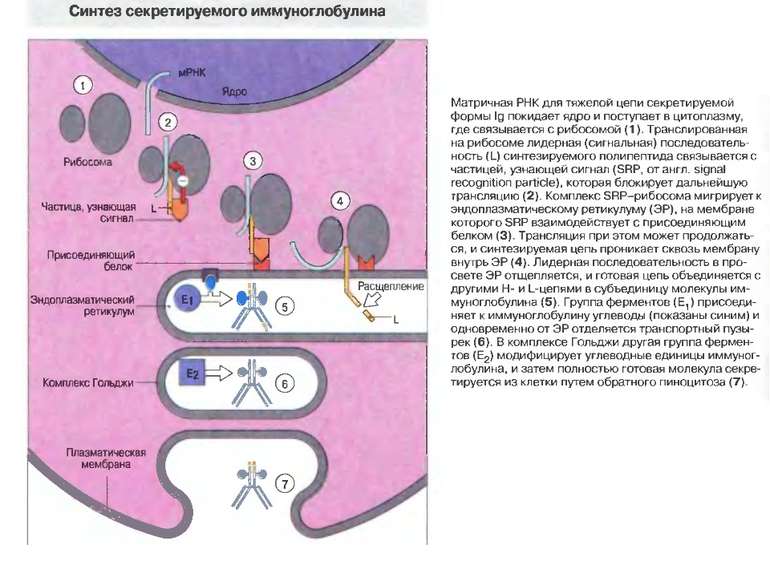
Синтез і транспорт Ig М у мембрани й у позаклітинний простір: 1-1- цис-; 2 - транс- сторони апарата Гольджі
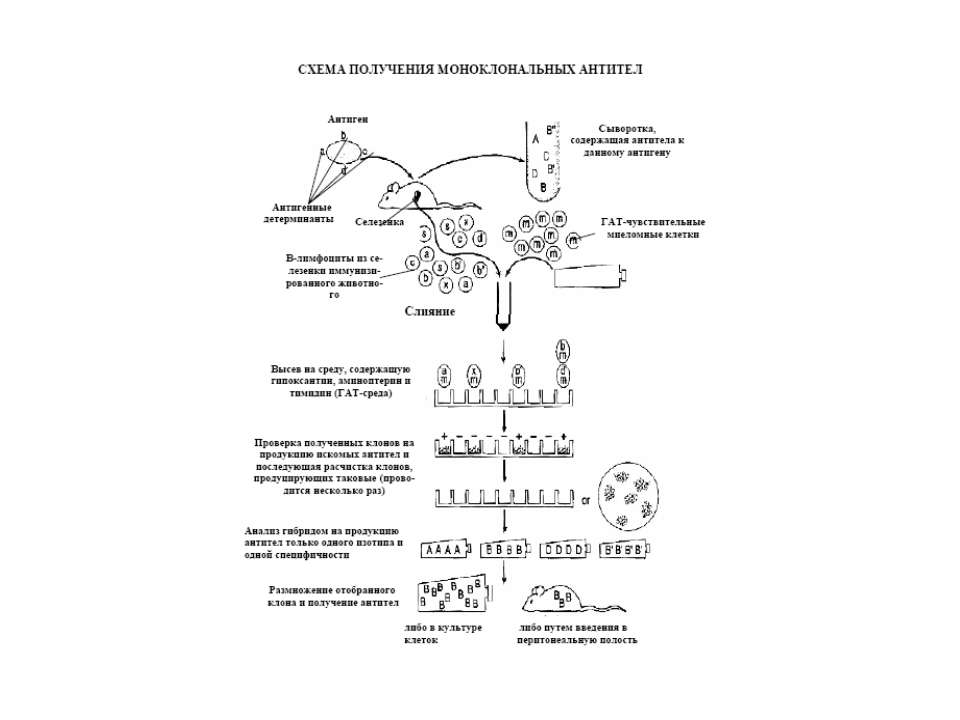
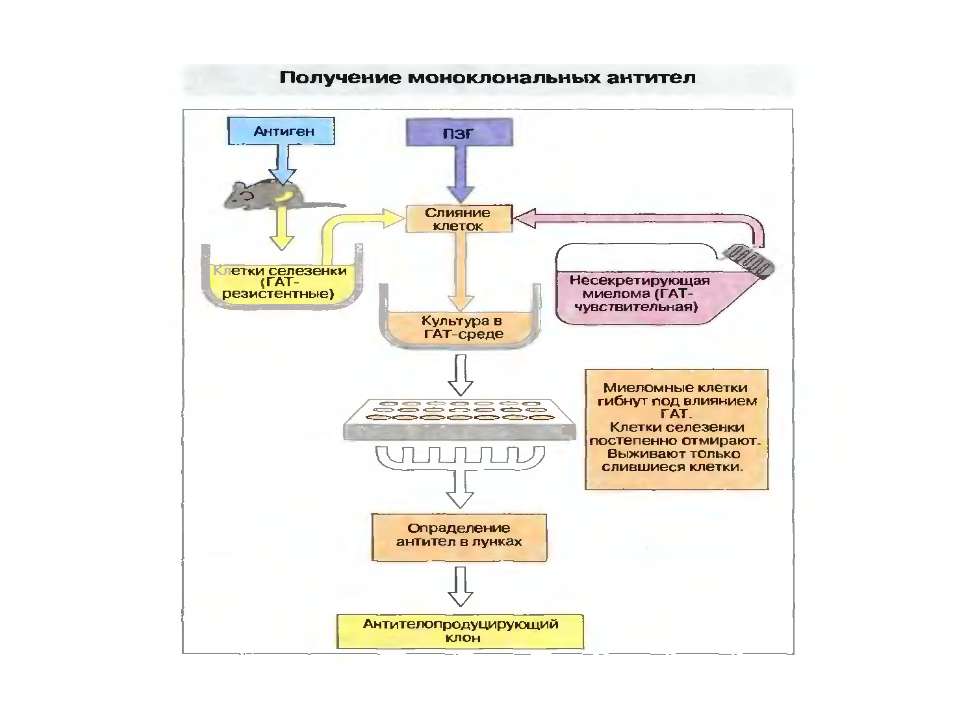
Схожі презентації
Категорії











