Презентація на тему:
частные аспекты метаболизма углеводов
Завантажити презентацію


































частные аспекты метаболизма углеводов
Завантажити презентаціюПрезентація по слайдам:

Енергетичні зв’язки між анаболізмом і катаболізмом

Стадії перетворення основних поживних речовин клітини

Важные метаболические пути углеводов Синтез/распад крахмала Синтез распад гликогена Глюконеогенез С4 путь фотосинтеза Фотосинтез по типу толстянковых Фотодыхание

Шляхи утилізації пірувату

Метаболизм гликогена Гликоген служит в животном организме резервом углеводов, из которого по мере метаболической потребности могут высвобождаться глюкозофосфат или глюкоза. Хранение в организме собственно глюкозы неприемлемо из-за ее высокой растворимости: высокие концентрации глюкозы создают в клетке высоко гипертоническую среду, что приводит к притоку воды. Напротив, нерастворимый гликоген осмотически почти неактивен. Брожение

Метаболизм гликогена Гликоген животных, как и амилопектин растений, представляет собой разветвленный гомополимер глюкозы, в котором остатки глюкозы соединены α(1→4)-гликозидной связью. Связи в точках ветвления находятся в положении α(1→6) примерно каждого 10-го остатка. Таким образом, возникает древовидная структура с молекулярной массой >1 107 Да (до 50 000 остатков), в которой имеется только одна свободная аномерная ОН-группа, т. е. только один восстанавливающий конец. Брожение

Метаболизм гликогена

Анаболизм гликогена Гликоген печени никогда не расщепляется полностью. Как правило, укорачиваются или удлиняются (при высоком содержании глюкозы) только невосстанавливающие концы древовидной структуры. Удлинение цепи катализируется гликоген-синтазой [2]. Так как образование гликозидных связей между сахарами является эндоэргической реакцией, вначале в реакции глюкозо-1-фосфата с уридинтрифосфатом [УТФ (UTP)] образуется активированный предшественник — УДФ-глюкоза (UDP-глюкоза) [1]

Анаболизм гликогена После этого остаток глюкозы легко переносится с этого промежуточного соединения на гликоген. Когда растущая цепь достигает определенной длины (>11 остатков), специальный фермент ветвления гликогена (1,4→1,6-трансгликозидаза ) [3] катализирует перенос концевого олигосахарида, состоящего из 6-7 остатков, на 6-ОН остаток глюкозы той же или другой цепи гликогена с образованием точки ветвления [α(1→6)-связи] Дальнейшее удлинение этого фрагмента осуществляется гликоген-синтазой, образующей α(1→4)-связи.

Катаболизм гликогена Разветвленная структура гликогена облегчает быстрое освобождение углеводных остатков. Наиболее важным ферментом деградации гликогена является гликоген-фосфорилаза [4], отщепляющая от невосстанавливающего конца цепи остатки глюкозы в виде глюкозо-1-фосфата. Чем больше таких концов, тем больше молекул фосфорилазы могут действовать одновременно. Образование глюкозо-1-фосфата вместо глюкозы имеет то преимущество, что для включения освобожденных остатков глюкозы в гликолиз или ГМП не требуется АТФ. .

Катаболизм гликогена Благодаря структуре гликоген-фосфорилазы, процесс последовательного отщепления останавливается за 4 остатка глюкозы от точки разветвления. Точки ветвления удаляются двумя другими ферментами [5 и 6]. Вначале трисахарид боковой цепи переносится [5] к невосстанавливающему концу главной цепи. Затем 1,6-гликозидаза [6] отщепляет остающийся единичный остаток глюкозы в точке ветвления в виде свободной глюкозы, после чего неразветвленная цепь, может вновь расщепляться фосфорилазой. .

Баланс гликогена В организме человека может содержаться до 450 г гликогена, треть из которого накапливается в печени, а остальное — главным образом в мышцах. Содержание гликогена в других органах незначительно. Гликоген печени служит прежде всего для поддержания уровня глюкозы в крови в фазе пострезорбции. Поэтому содержание гликогена в печени варьирует в широких пределах. При длительном голодании оно падает почти до нуля, после чего начинается снабжение организма глюкозой с помощью глюконеогенеза .

Баланс гликогена Гликоген мышц служит резервом энергии и не участвует в регуляции уровня глюкозы в крови. В мышцах отсутствует глюкозо-6-фосфатаза, поэтому гликоген мышц не может быть источником глюкозы в крови. По этой причине колебания содержания гликогена в мышцах меньше, чем в печени.

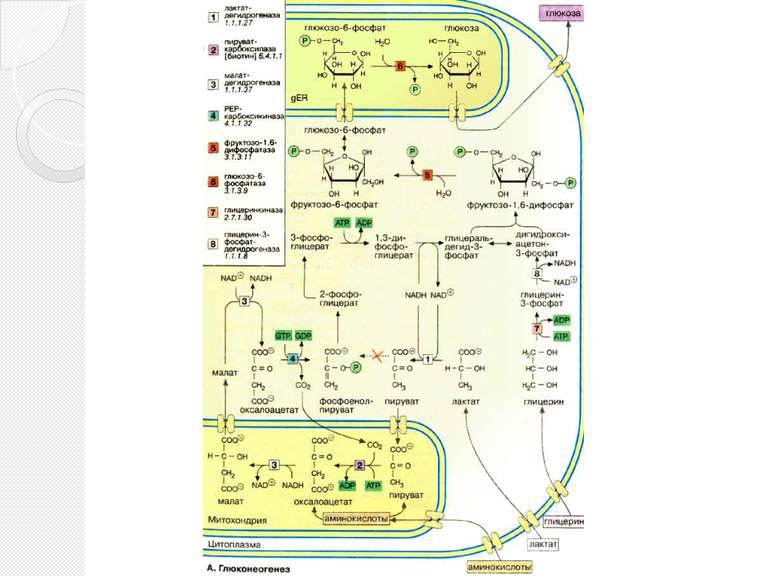
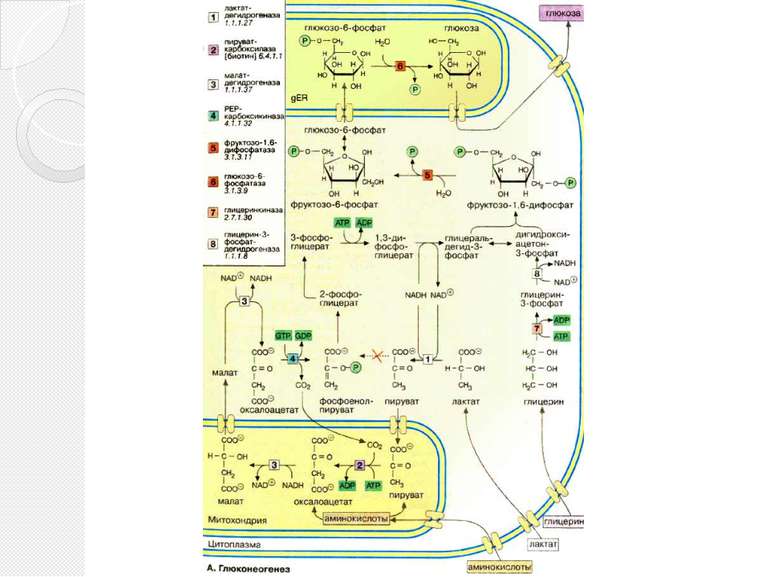
Глюконеогенез Некоторые ткани, такие, как мозг и эритроциты, зависят от постоянного снабжения глюкозой. Если получаемое с пищей количество углеводов недостаточно, необходимая концентрация глюкозы в крови может поддерживаться некоторое время за счет расщепления гликогена печенью. Если истощены также и эти запасы, в печени запускается синтез глюкозы de novo, глюконеогенез. Наряду с печенью высокой глюконеогенезной активностью обладают также клетки почечных канальцев. .

Глюконеогенез Исходными соединениями в глюконеогенезе являются аминокислоты мышечной ткани. При длительном голодании это приводит к массивному распаду мышечного белка. Другими важными исходными веществами для синтеза глюкозы служат лактат, образующийся в эритроцитах и мышечной ткани при недостатке О2 , а также глицерин, образующийся при расщеплении жиров. Напротив, жирные кислоты не могут трансформироваться в глюкозу в организме животных, так как в данном случае деградация жирных кислот не является анаплеротическим процессом . В организме человека за счет глюконеогенеза образуется несколько сотен граммов глюкозы в сутки.


Глюконеогенез В то время как гликолиз протекает в цитоплазме, глюконеогенез происходит также в митохондриях и эндоплазматическом ретикулуме. Первые стадии реакционной цепи протекают в митохондриях. Причиной такого «обходного» пути является неблагоприятная константа равновесия пируваткиназной реакции (см. с. 152). Для перевода пирувата непосредственно в фосфоенолпируват(PEP) недостаточно энергии расщепления АТФ. Пируват, образующийся из лактата или аминокислот, переносится в матрикс митохондрий и там карбоксилируется в оксалоацетат в биотинзависимой реакции, катализируемой пируваткарбоксилазой [2]. Оксалоацетат является промежуточным метаболитом цитратного цикла. Поэтому аминокислоты, которые включаются в цитратный цикл или конвертируются в пируват, могут непосредственно превращаться в глюкозу (глюкогенные аминокислоты,).

Глюконеогенез Оксалоацетат, образующийся в митохондриальном матриксе, восстанавливается в малат [3]. который может переноситься в цитоплазму с помощью специальных переносчиков. Оксалоацетат может также переноситься из митохондрии в цитоплазму после переаминирования в аспартат. В цитоплазме малат вновь превращается цитоплазматической малатдегидрогеназой в оксалоацетат, который в реакции, катализируемой ГТФ-зависимой РЕР-карбоксикиназой [4], переводится в фосфоенолпируват. Последующие стадии до фруктозо-1,6-дифосфата представляют собой модификации соответствующих реакций гликолиза. При этом для образования 1,3-дифосфоглицерата дополнительно расходуется АТФ

Гликолиз
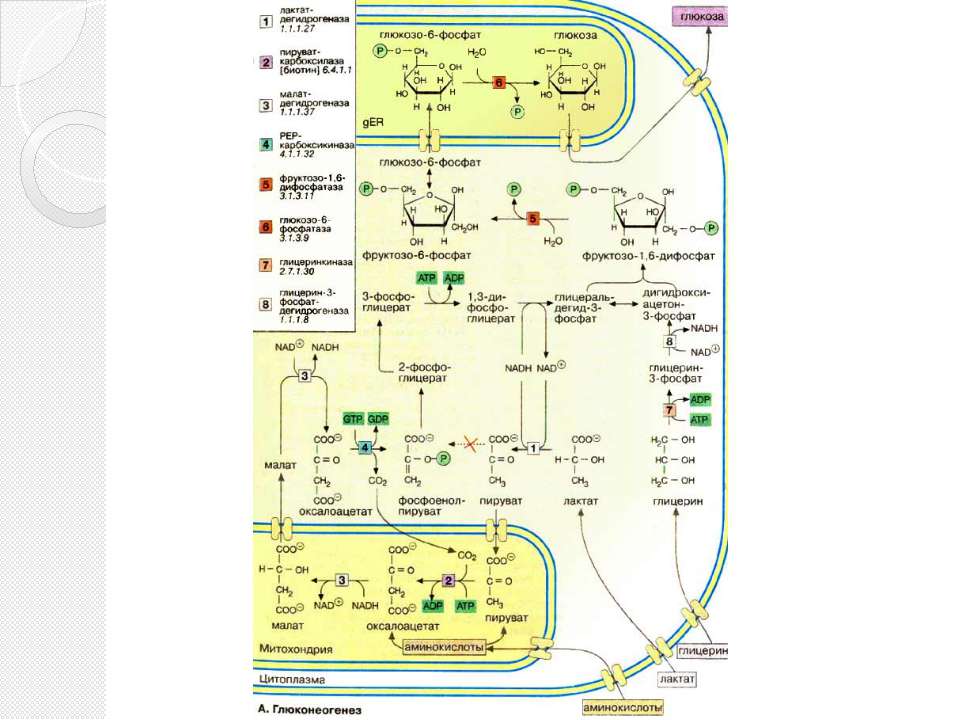

Глюконеогенез Две глюконеогенез-специфичные фосфатазы отщепляют по очереди фосфатные остатки от фруктозо-1,6-дифосфата. Промежуточной стадией является изомеризация фруктозо-6-фосфата в глюкозо-6-фосфат, одна из реакций гликолиза. Глюкозо-6-фосфатаза печени [5] является мембранным ферментом, локализованным внутри гладкого эндоплазматического ретикулума. Перенос глюкозо-6-фосфата в эндоплазматический ретикулум и возврат образующейся глюкозы в цитоплазму осуществляется специфическими переносчиками. Из цитоплазмы глюкоза поступает в кровь. Глицерин прежде всего фосфорилируется [7] в положении 3. Образующийся 3-глицерофосфат окисляется НАД+-зависимой дегидрогеназой [8] в дигидроксиацетон-3-фосфат, который далее включается в глюконеогенез.

Цикл Хэтча-Слэка-Карпилова

Цикл Хэтча-Слэка-Карпилова

Цикл Хэтча-Слэка-Карпилова

Цикл Хэтча-Слэка-Карпилова

Цикл Хэтча-Слэка-Карпилова

Цикл Хэтча-Слэка-Карпилова

Цикл Хэтча-Слэка-Карпилова

Фотосинтез по типу толстянковых

Фотодыхание

Фотодыхание

Фотодыхание

Фотодыхание

Фотодыхание

Фотодыхание
Схожі презентації
Категорії











