Презентація на тему:
класифікація
Завантажити презентацію



 ...")
 фотосинтезу світлова (світлозалежна) стадія – фотофізичні (погл...")













 — широко распространённый в п...")















класифікація
Завантажити презентаціюПрезентація по слайдам:

Трофічна класифікація живих організмів Різноманіття способів живлення є характерним для прокаріотичних організмів. Натомність всі рослини - фотолітоавтотрофи, всі тварини і гриби - хемоорганогетеротрофи

Рослини – основне джерело біологічної речовини В процесі ф/с наземні рослини утворюють 100-170 млрд. т, а рослини океанів — 60-70 млр. т біомаси в рік (у перерахунку на суху речовину). Загальна маса рослин на Землі дорівнює 2 400 млр. т (90%-це целюлоза). Загальна маса тварин і м/о - 23 млрд. т, що становить коло 1% від рослинної біомаси.
 ...")
Фотосинтез (від грец. φωτο- — світло та грец. σύνθεσις — синтез, сукупність) — процес синтезу органічних сполук з неорганічних (вуглекислого газу та води) з використанням енергії світла: відбувається за участю фотосинтетичних пігментів: (хлорофіл у рослин, хлорофіл, бактеріохлорофіл і бактеріородопсин у бактерій), часто з виділенням кисню (оксигеннний фотосинтез) як побічного продукту. включає довгу послідовність координованих хімічних реакцій.
 фотосинтезу світлова (світлозалежна) стадія – фотофізичні (погл...")
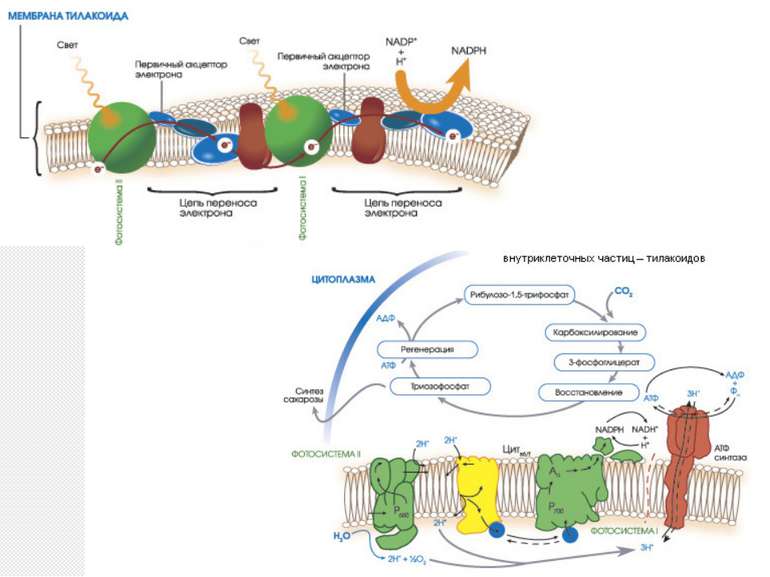
Фази (стадії) фотосинтезу світлова (світлозалежна) стадія – фотофізичні (поглинання сввітлових квантів пігментами, їх перехід в збуджений стан і передача енергії до інших молекул фотосистеми) та фотохімічні (розділення зарядів в реакційному центрі, перенесення електронів по фотосинтетичному електронотранспортному ланцюзі, що закінчується синтезом АТФ і НАДФН) реакції темнова стадія – зв’язування СО2 та синтез органічних речовин з використанням енергії, накопиченої на світлозалежній стадії (цикл Кальвіна).

Загальне рівняння фотосинтезу 6СО2 + 6Н2О С6Н12О6 + 6О2 світло

Хеміосмотична теорія Мітчела Енергія, що вивільнюється внаслідок роботи електрон-транспортного ланцюга, спочатку накопичується у формі трансмембранного електрохімічного градієнта іонів водню (DmН+) озрядка накопиченого DmН+ відбувається за участю локалізованого в тій самійц мембрані АТФ-синтазного комплексу, Н+ повертається по градієнту через Н+-АТФ-синтазу, при цьому без утворення будь-яких високоенергетичних інтермедіантів з АДФ та неорганічного фосфата утворюється АТФ. Спрягаючі мембрани в інтактному стані непроникні для будь-яких іонів

Локалізація фотосинтетичних процесів процеси світлової стадії фотосинтезу відбуваються на мембранах тилакоїдів, які є складовою частиною хлоропластів процеси темнової стадії відбуваються в стромі хлоропластів

Світлозбираючи комплекси містять набір пігментів – хлорофіли, каротиноїди та фікобіліни

Світлозбираючи комплекси містять набір пігментів – хлорофіли, каротиноїди та фікобіліни

Спектри поглинання деяких фотосинтетичних пігментів

ХЛОРОФІЛИ

Хлорофіл Хлорофилл является хорошим сенсибилизатором — легко возбуждается при поглощении света и обладает способностью передавать энергию (служить донором энергии) другим молекулам (акцепторам энергии). Передача энергии возможна лишь при условии, что спектр флуоресценции пигмента-донора перекрывается со спектром поглощения пигмента-акцептора. Таким путем происходит резонансный перенос энергии возбуждения пигментами в хлоропластах от молекулы донора к молекуле акцептора.

Каротиноїди

Фікобіліни

енергетичні стани молекули хлорофіла молекула хлорофіла здатна знаходитись в тріплетному збудженому стані протягом часу, достатнього для здійснення хімчної роботи

Z-схема фотосинтеза

Z-схема фотосинтеза
 — широко распространённый в п...")
Никотинамидадениндинуклеотидфосфат (НАДФ, NADP) — широко распространённый в природе кофермент некоторых дегидрогеназ — ферментов, катализирующих окислительно-восстановительные реакции в живых клетках. NADP принимает на себя водород и электроны окисляемого соединения и передает их на другие вещества. В хлоропластах растительных клеток NADP восстанавливается при световых реакциях фотосинтеза и затем обеспечивает водородом синтез углеводов при темновых реакциях. NADP, — кофермент, отличающийся от NAD содержанием ещё одного остатка фосфорной кислоты, присоединённого к гидроксилу одного из остатков D-рибозы, обнаружен во всех типах клеток.

Фотосистеми

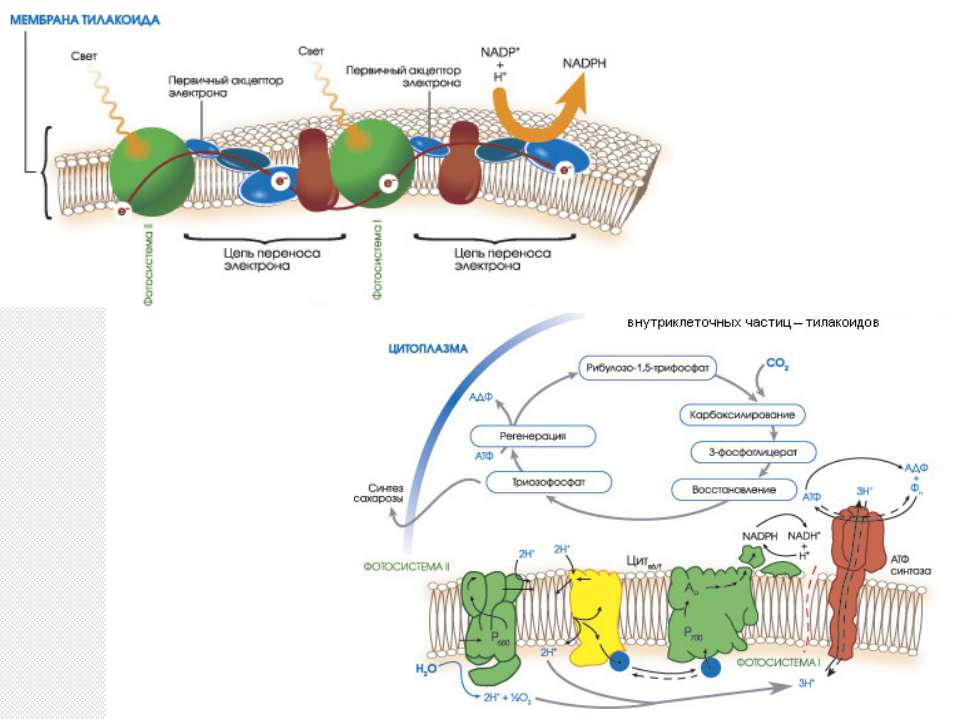
Фотосистема ІІ Фотосистема — сукупність СЗК, фотохімічного реакційного центру і переносників електрона. Світлозбиральний комплекс II може містити до 200 молекул хлорофілу а, 100 молекул хлорофілу b, 50 молекул каротиноїдів і 2 молекули феофітину. Реакційний центр фотосистеми II є пігмент-білковим комплексом, розташованим в тилакоїдних мембранах і оточеним СЗК. У ньому знаходиться димер хлорофілу а з максимумом поглинання при 680 нм (П680). На нього кінець-кінцем передається енергія кванта світла з СЗК, внаслідок чого один з електронів переходить у вищий енергетичний стан, зв'язок його з ядром ослаблюється, і збуджена молекула П680 стає сильним відновником (E0=-0,7 В).

Фотосистема ІІ П680 відновлює феофітин, надалі електрон переноситься на хинони, що входять до складу ФС II, і далі на пластохинони, транспортується у відновленій формі до b6f комплексу. Одна молекула пластохинона переносить 2 електрони і 2 протони, які беруться зі строми. Заповнення електронної вакансії в молекулі П680 відбувається за рахунок води. До складу ФС II входить водоокислюючий комплекс, що містить в активному центрі іони марганцю в кількості 4 штук. Для утворення однієї молекули кисню потрібно дві молекули води, що дають 4 електрони. Тому процес проводиться в 4 такти, і для його повного здійснення потрібно 4 кванти світла. Комплекс знаходиться з боку внутрішньотилакоїдного простору, і отримані 4 протони викидаються в нього. Таким чином, сумарний результат роботи ФС II — це окислення 2 молекул води за допомогою 4 квантів світла з утворенням 4 протонів у внутрішньотилакоїдному просторі і 2 відновлених пластохинонів в мембрані.

Будова кисеньвиділяючого центру фотосистеми ІІ

b6f або b/f-комплекс b6f комплекс є насосом, що перекачує протони із строми у внутрішньотилакоїдний простір і створює градієнт їхньої концентрації за рахунок енергії, що виділяється в окисно-відновних реакціях електронно-транспортного ланцюжка. 2 пластохинони забезпечують перекачування 4 протонів. Надалі трансмембранний протонний градієнт (pH строми, близький до 8, внутрішньотилакоїдного простору — 5) використовується для синтезу АТФ трансмембранним ферментом АТФ-синтазою.

Фотосистема І Світлозбиральний комплекс I містить приблизно 200 молекул хлорофілу. У реакційному центрі першої фотосистеми знаходиться димер хлорофілу а з максимумом поглинання при 700 нм (П700). Після збудження квантом світла він відновлює первинний акцептор — хлорофіл а, той — вторинний (вітамін K1), після чого електрон передається на філохінон, від нього на феридоксин, який і відновлює НАДФ за допомогою ферменту феридоксин-НАДФ-редуктази. Білок пластоціанін, окислений в b6f комплексі, транспортується до реакційного центру першої фотосистеми з боку внутрішньотилакоїдного простору й відновлює окислений П700.

Транспорт електронів і протонів в мембрані тилакоїдів

Циклічний і псевдоциклічний транспорт електрона Крім повного нециклічного шляху електрона, описаного вище, виявлено циклічний і псевдоциклічний. Суть циклічного шляху полягає в тому, що феридоксин замість НАДФ відновлює пластохинон, який переносить його назад на b6f комплекс. В результаті утворюється більший протонний градієнт і більше АТФ, але не виникає НАДФН. При псевдоциклічному шляху феридоксин відновлює кисень, який надалі перетворюється на воду і може бути використаний у фотосистемі II. При цьому також не утворюється НАДФН.

Модель будови АТФ-синтази

ЦИКЛ КАЛЬВІНА

Основні стадії циклу Кальвіна Карбоксилирование Карбоксилирование рибулозо-1,5-бисфосфата (5-углеродное соединение) осуществляется рубиско в несколько стадий. На первой кетонная группа рибулозы восстанавливается до спиртовой, между 2 и 3 атомами углерода устанавливается двойная связь. Полученное соединение нестабильно и именно оно карбоксилируется с образованием 2-карбокси-3-кето-D-арабитол-1,5-бисфосфата. Его структурный аналог 2-карбокси-D-арабитол-1,5-бисфосфат ингибирует весь процесс. Новое, уже 6-углеродное соединение, также нестабильно и распадается на две молекулы 3-фосфоглицериновой кислоты (3-фосфоглицерат, 3-ФГА).

Основні стадії циклу Кальвіна Восстановление Восстановление 3-фосфоглицериновой кислоты (3-ФГА) происходит в две реакции. Сначала каждая 3-ФГА с помощью 3-фосфоглицераткиназы и с затратой одной АТФ фосфорилируется, образуя 1,3-бисфосфоглицериновая кислота (глицерат-1,3-бисфосфат). Затем под действием глицеральдегид-1,3-фосфатдегидрогеназы бисфосфоглицериновая кислота восстанавливается НАД(Ф)·H (у растений и цианобактерий; у пурпурных и зелёных бактерий восстановителем является НАД·H) параллельно с отщеплением одного остатка фосфорной кислоты. Образуется глицеральдегид-3-фосфат (фосфоглицеральдегид, ФГА, триозофосфат). Обе реакции обратимы.

Основні стадії циклу Кальвіна Регенерация На последней стадии 5 молекул глицеральдегид-3-фосфатов превращаются в три молекулы рибулозо-1,5-бисфосфата. Вначале под действием трифосфат-изомеразы глицеральдегид-3-фосфат изомеризуется в дигидроксиацетон-фосфат. Фруктозабисфосфат-альдолаза объединяет их в фруктозо-6-фосфат с отщеплением остатка фосфорной кислоты. Затем следует ряд реакций перестройки углеродных скелетов и образуется рибулозо-5-фосфат. Он фосфорилируется фосфорибулокиназой и рибулозо-1,5-бисфосфат регенерируется.

ЦИКЛ КАЛЬВІНА

Принципова подібність процесів хеміосмотичного спряження
Схожі презентації











Категорії











