Презентація на тему:
ФІЗИЧНІ ОСНОВИ ФУНКЦІОНУВАННЯ БІОЛОГІЧНИХ МЕМБРАН
Завантажити презентацію






.Осно...")











 припущенням про...")


 мембрани становить подвійний ліпідний ...")





 або (3.4) Рівняння Фіка описує один із найпоширеніших механізмів пасивн...")


 (3.8) k-коефіцієнт розподілу, показує, яку частину концентрація всереди...")

: (3.12) де - швидкість переносу вод...")












ФІЗИЧНІ ОСНОВИ ФУНКЦІОНУВАННЯ БІОЛОГІЧНИХ МЕМБРАН
Завантажити презентаціюПрезентація по слайдам:

ФІЗИЧНІ ОСНОВИ ФУНКЦІОНУВАННЯ БІОЛОГІЧНИХ МЕМБРАН

1 Молекулярна організація мембран 1.1 З історії фізики мембран. 1.2 Структура і властивості і функції мембранних ліпідів. 1.3 Структура, властивості і функції мембранних білків. 1.4 Структура, властивості і функції мембранних вуглеводів. 4.5 Основні види взаємодій, які формують мембрани. Моделі мембран. 2 Транспорт речовин через мембрани 2.1 Дифузія нейтральних молекул. 2.2 Осмос. 2.3 Фільтрація. 2.4 Дифузія іонів через мембрану 2.5 Полегшена дифузія. 2.6 Активний транспорт.

ФІЗИЧНІ ОСНОВИ ФУНКЦІОНУВАННЯ БІОЛОГІЧНИХ МЕМБРАН 1 Молекулярна організація мембран 1.1 З історії фізики мембран. Формує клітину і підтримує чітко визначений внутрішньо клітинний гомеостаз плазматична, або поверхнева, мембрана. Вона вибірково регулює швидкість руху речовин у клітину та з клітини за допомогою транспортних систем, до складу яких входять специфічні переносники і мембранозв'язані ферменти. Плазматична мемб рана є складним ферментативним, постійно функціонуючим комплексом, що забезпечує такі прояви життєдіяльності, як дихання, імунітет, травлення, міжклітинні взаємодії, передачу збудження тощо. Вона перша зустрічає, класифікує впливи зовнішнього середовища і трансформує їхню енергію біологічного збудження.

Вчені вважають, що ідея про існування особливої перегородки, яка оточує клітину, вперше була висловлена Клодом Бернаром. Вивчаючи проникність барвників у клітину, Карл Негіллі у 1855 р. також висловив думку про те, що зовні протоплазма вкрита особливою оболонкою, яка виконує функцію бар'єру проникності. І тільки через три десятиріччя, повторюючи досліди Негіллі і проводячи власні оригінальні дослідження, Пфеффер встановив, що клітина поводить себе як мініатюрний осмометр, змінюючи ступінь набухання залежно від концентрації зовнішнього розчину. На підставі своїх експериментів та ідей Бернара і Негіллі Пфеффер у 1890 р. сформулював поняття клітинної (плазматичної) мембрани як специфічного утвору, що регулює надходження у клітину і вихід з неї різних речовин. Уже в ті часи вчених хвилювало питання про хімічний склад і молекулярну організацію мембран. Першими можна вважати дослідження Овертона (1895-1899), в яких він вивчав проникнення речо вин різної хімічної природи у клітину. Результати його досліджень показали, що в клітину швидше проникають ті речовини, які краще розчиняються у жирах. Виходячи з цього, а також з пфефферівського визначення поверхневої мембрани як бар'єру про никності, Овертон у 1902 р. висловив припущення, що мембрана повністю або частково побудована з ліпідів. Отже, в кінці минулого і на початку нашого століття, не тільки було встановлено існування плазматичної мембрани, а й ідентифікований один з її основних хімічних компонентів.

У 1925 р. Гортер і Грендаль екстрагували ліпіди з еритроцитів і порівнювали площу моношару цих ліпідів з площею клітин. Автори встановили, що площа моношару екстрагованих ліпідів достатня щоб вкрити еритроцит двічі. Крім цього, Гортер Грендель, мабуть, вперше висловили думку про те, що такий подвійний шар ліпідів певним чином зв’язаний з білками. У 1925 р. вийшла робота Фріке, в якій не тільки повністю підтримувалася ідея існування мембрани, а й було виміряно її товщину. Фріке встановив, що ємність мембран еритроцитів становить , що відповідає товщині діелектрика в 3,3 нм. Таким діелектриком на поверхні клітини може бути лише подвійний шар ліпідів.
.Осно...")
Завершенням цього етапу досліджень стали роботи Даніелі і Давсона (1935).Основний зміст цих досліджень полягав у вимірюванні поверхневого натягу на межі розділення масла і водних розчинів. Автори встановили, що поверхневий натяг краплі ліпідів на межі поділу ліпід-вода набагато перевищував поверхневий натяг на межі поділу ліпід-цитоплазма. Даніелі і Давсон припустили, що зниження поверхневого натягу може бути зумовлене речовинами протоплазми, передусім білками. Ці дані з урахуванням попередніх робіт дали змогу вченим висловити гіпотезу про будову біологічних мембран. За цією гіпотезою, плазматична мембрана складається з подвійного ліпідного шару, вкритого з внутрішнього і зовнішнього боків моношарами білків. Отже, існування на поверхні клітин плазматичної мембрани постулювалось не тільки на підставі даних проникнення у клітину різних речовин, а й на основі електричного опору і ємності, що притаманні поверхневим шарам клітин. Проте фактичні докази існування мембрани було добуто після широкого впровадження у біологію методів електронної мікроскопії. Так вдалося «побачити» мембрану, про існування якої біологи здогадувалися. З 1955р. почали виявляти внутрішньоклітинні мембрани.

На електронно-мікроскопічних фотографіях мембрана має вигляд плівки до 10 нм завтовшки, яка складається з трьох шарів: одного (середнього) світлого і двох темних, що прилягають до першого (рис.3.1). Плазматична мембрана асиметрична: з внутрішнього цитоплазматичного боку електронно-щільний шар товстіший і темніший, ніж зовні. Отже, мембрани є місцем розмежування вмісту клітин від зовнішнього середовища і поділу внутрішнього простору клітин на відсіки, або компартменти. Компартменталізація зіграла важ ливу роль як в добіологічній, так і в біологічній еволюції. Існування живого як відкритої системи неможливе без контакту із зовнішнім середовищем. У зв'язку з цим мембрани не тільки розмежовують, а й здійснюють регуляцію взаємозв'язків клітин і зовнішнього середовища, взаємозв'язків між окремими клітинами і компартментами клітин. Такі взаємозв'язки базуються на обміні речовиною, енергією та інформацією. Тепер твердо встановлено, що біологічні мембрани - це складні надмолекулярні структури, побудовані з ліпідів, білків, вуглеводів і неорганічних іонів. До складу мембран входить і вода. Між молекулами, ліпідів, білків і води існує декілька типів взаємодій, які і визначають структуру біологічних мембран. Для того, щоб зрозуміти як формуються мембрани і які їх властивості треба знати склад і будову молекул які утворюють мембрани.

1.2 Структура, властивості і функції мембранних ліпідів. За хімічною природою мембранні ліпіди можна віднести до трьох класів: фосфоліпіди, гліколіпіди і нейтральні ліпіди. В середньому ліпіди становлять близько 40% сухої маси біологічних мембран, з яких 80% належить фосфоліпідам. До складу ліпідів входять жирні кислоти, спирти, альде гіди, азотисті основи, вуглеводи, фосфорна кислота тощо. Ці компоненти ліпідів зв'язуються ефірними, амідними, фосфодиефірними та іншими типами зв'язків. Жирні кислоти - один з основних компонентів ліпідів мембран; вони певною мірою зумовлюють хімічні властивості ліпідів, взаємодію між молекулами ліпідів і білків, що в кінце вому результаті визначає структурні особливості самих мембран і їхню ферментативну активність. Жирні кислоти, що входять до складу ліпідів мембран вищих організмів, мають ряд спільних ознак. Майже всі вони мають парне число атомів вуглецю (від 14 до 22, найчастіше 16 або 18).Вміст ненасичених жирних кислот, як правило, вищий, ніж насичених.

Вуглеводневі ланцюги жирних кислот зумовлюють гідрофобні властивості мембранних ліпідів. Проте якби ліпіди характеризувались тільки гідрофобними властивостями, вони були б не здатні до утворення мембранних структур. Молекуи мембранних ліпідів характеризуються і різною мірою вираженою гідрофільністю. Гідрофільні властивості ліпідних молекул зумовлюються полярними головками, які можуть мати різну хімічну будову. В молекулах фосфоліпідів полярні головки утворенні похідними фосфорної кислоти, а в молекулах гліколіпідів – похідними цукрів. Отже, молекули мембранних ліпідів характеризуються наявністю двох кінців: полярної (гідрофільної)головки, що утворює з водою водневі, або дипольні, зв’язки, і неполярного хвоста (гідрофобного) (рис.3.2.). У більшості мембранних ліпідів головки мають одночасно позитивний і негативний заряди.

¾ ¼

1.3 Структура, властивості і функції мембранних білків. Хімічні властивості мембранних білків вивчено значно гірше, ніж ліпідів. Це пов’язано з тим, що мембранні білки нерозчинні у воді і погано розчинні в органічних розчинниках. Оскільки білки в мембрані перебувають у контакті з ліпідами, дуже важко встановити , чи справді білки в мембрані існують у такому стані, в якому їх виділяють. У зв’язку з цим для з’ясування нативної (або близької до неї) структури мембранних білків часто застосовують оптичні методи дослідження (круговий дихроїзм, інфрачервона спектроскопія, дисперсія оптичного повертання, парамагнітні зонди тощо).За допомогою цих методів встановлено, що в мембранах поряд з глобулярними білками присутні й фібрилярні білки, що мають α- та β- конфігурацію.

Мембранні білки за функцією поділяють на три групи: структурні білки, білки-ферменти та рецепторні білки. Проте сьогодні не можна однозначно визначити, скільки і які білки входять до складу тієї або іншої мембрани. Це пов'язано з труднощами виді лення білків у чистому вигляді, а також у можливості врахування асоціації і дисоціації поліпептидних ланцюгів у процесі виділення. Дуже можливо, що ліпіди і білки в мембранах зв'язані за рахунок гідрофобних ділянок молекул, а білок і вода - за рахунок гідрофільних. У зв'язку з цим не останнє місце у вивченні мембранних білків і розумінні їхньої поведінки має амінокислотний склад, оскільки тільки через амінокислоти реалізується зв'язок білків з іншими компонентами мембран. Якщо на поверхні молекули білка більше гідрофільних груп, то реалізуватимуться зв'язки з водою, що може привести до витягування молекул білка. Коли ж на поверхні молекули переважають гідрофобні групи, більшою мірою відбуватиметься гідрофобна взаємодія білкових і ліпідних молекул, що має значення в утворенні білково-ліпідних комплексів мембран.

1.4 Структура, властивості і функції мембранних вуглеводів. Наявність вуглеводних компонентів у мембра ні до останнього часу мало цікавила дослідників. Серед вуглеводів, що входять до складу мембран тварин них клітин, найчастіше зустрічаються полісахариди. Встановлено, що мукополісахариди зв'язані з ліпідами мембран і певною мірою відповідальні за антигенність клітин. Крім того, полісахаридам приписують також важливу роль у процесах адгезії клітин. Експериментально показано, що поверх ня клітин вкрита тонким клейким шаром. Він складається в основ ному з білків і полісахаридів. Оскільки клітинна поверхня рухлива, мембрана весь час деформується. Ці рухи змінюють розміщення макромолекул або поліпептидних ланцюгів, що й зумовлює зміну здатності клітинної поверхні до адгезії.

Встановлено, що за хімічним складом поверхня плазматичної мембрани тваринних клітин має щонайменше три основних компоненти: вуглеводні компоненти гліколіпідів, глікопротеїнів, кислі мукополісахариди. Кислі мукополісахариди дуже гідратовані і мають желеподібну форму, липкі або слизькі речовини, є міжклітинним "мастилом" і одночасно лабільним цементуючим матеріалом.

1.5 Основні види взаємодій, які формують мембрани. Моделі мембран. Виходячи з хімічного складу та асиметричної локалізації компонентів мембран можна сказати, що всі вони формують надмолекулярний мембранний комплекс за рахунок взаємодії і, передусім, за рахунок двох найважливіших типів нековалентних взаємодій: гідрофобних і гідрофільних. Гідрофобні взаємодії є результатом відштовхування води ліпофільними групами молекул. Оскільки молекули води мають більшу спорідненість до інших молекул води або полярних груп, то поблизу гідрофобних ділянок моле кул збільшується квазикристалічний порядок у структурі води. В разі відштовхування гідрофобних груп з водного оточення відбувається зменшення впорядкованості молекул води. Окрім гідрофобних і гідрофільних взаємодій важливу роль у структурі мембран відіграють іонні взаємодії і зв'язки через двовалентні іони, а також водневі зв'язки. Іонні зв'язки існують між моле кулами, які мають протилежні електричні заряди, наприклад, між фосфатидилсерином і основними білками. Двовалентні катіони, насамперед іони кальцію, можуть зв'язувати або дві молекули фосфоліпідів, або молекулу білка з молекулою фосфоліпіду.

Водневі зв'язки також можуть відігравати важливу роль у білок-ліпідних взаємодіях в мембранах, оскільки багато груп мембранних білків і ліпідів можуть бути донорами або акцепторами водню. Застосування термодинамічного принципу мінімуму вільної енергії до моделей структури біологічних мембран вимагає виконання двох умов: 1) білки і ліпіди мають бути розміщені в мембранах таким чином, щоб максимально можливе число полярних груп перебувало в контакті з водою і іншими полярними групами; 2) неполярні вуглеводневі ланцюги ліпідів і амінокислотних за лишків мають розміщуватися так, щоб якнайменше контактувати з водою.

Гортер і Грендель перші припустили, що в основі структури мембрани лежить бімолекулярний шар ліпідів. В 1935 р. Даніелі і Даусон запропонували модель мембрани, яка протягом кількох десятиріч була основною в розумінні структури біологічних мембран. Згідно з моделлю Даніелі - Даусона серцевина мембрани, її внутрішня частина, складається з двох ліпідних моношарів, молекули яких розміщені паралельно одна одній і орієнтовані так, що неполярні "хвости" обох шарів спрямовані один до одного. Полярні кінці ліпідів спрямовані назовні по обидва боки мембрани. З обох боків такий бімолекулярний шар ліпідів покритий білками, які зв'язані з полярними кінцями ліпідів, завдяки електростатичним силам і захищають ліпіди від безпосереднього контакту з во дою. Подвійний шар ліпідів, згідно з цією моделлю, е основним енергетичним бар'єром для проникнення гідрофільних речовин.
 припущенням про...")
З огляду на це модель було модифіковано (Стейн.Даніелі, І956) припущенням про існування в структурі мембран полярних пор. Припускалось, що пори утворюються білками, які сполучають обидві поверхні мембран і зв'язані з ланцюгами ліпідів гідрофобни ми взаємодіями, Згідно з моделлю кількість білків, що проника ють крізь подвійний шар ліпідів, була обмежена. Основні принципи моделі Даніелі-Даусона були використані для створення концепції унітарної мембрани Робертсоном (І964). Згідно з цією концепцією, яка грунтувалась тільки на результатах електронно-мікроскопічних досліджень, в основі всіх мембран, незалежно від їхньої природи, складу і функції, лежить одна і та ж структура.

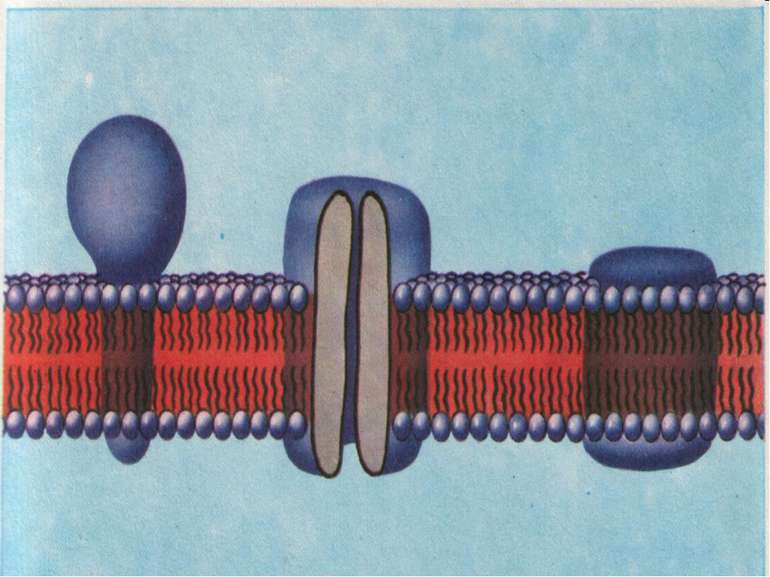
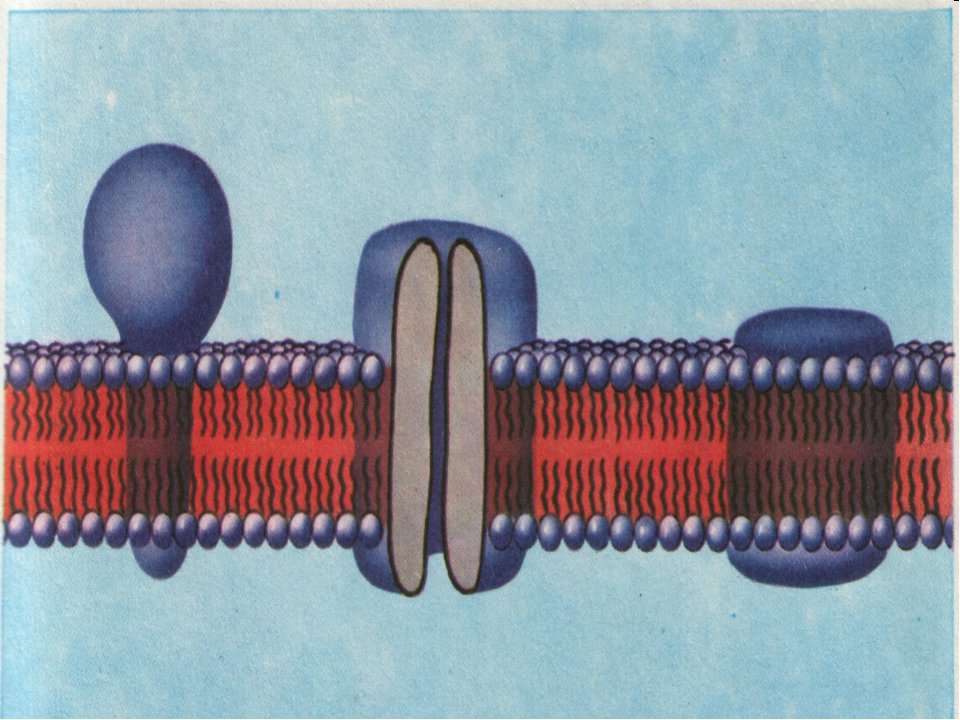
У 60-х роках на підставі даних електронної мікроскопії, реконструкції мембран, дифракції рентгенівського проміння з'я вились ряд моделей, в яких припускалась субодинична будова мемб ран. Пізніше ці моделі було відкрито, оскільки твердо встанов лено, що в основі мембран лежить не регулярна субодинична струк тура ліпідів, а ламелярний подвійний шар. Останнім часом досить перспективно вважається "рідинно-мозаїчна» модель, схема якої показана на рис.3

Рис.3.3.
 мембрани становить подвійний ліпідний ...")
Згідно з цією моделлю основу (матрицю) мембрани становить подвійний ліпідний шар. Для розуміння ролі білків в структурі мембрани, останні поділяються на дві групи: периферичні та інтегральні білки. Периферичними вважаються білки, слабо зв'язані з ліпідами (електростатичні сили) і які можуть легко виділятися з мембра ни при збільшенні іонної сили розчину, або дією хелатуючих агентів. Ці білки становлять близько 30% всіх мембранних білків. Інтегральні білки досить міцно зв'язані з ліпідами і часто виділяються з мембрани (під впливом органічних розчинників, детергентів, денатуруючих агентів тощо) разом з ліпідами.

Ці білки в чистому вигляді малорозчинні в нейтральних водних і буферних розчинах, мають аліфатичну при роду і входять до складу гліко- або ліпопротеїнів. Глобули гліко- і ліпопротеїнів занурені в ліпідні шари, деякі з них мають трансмембранну локалізацію. Чергування ділянок білків та ліпідів і дає "мозаїчну" картину мембрани. Більша частина фосфоліпідів представлена переривчастим біомолекулярним шаром, полярні групи яких перебувають у контак ті з водою або з білками. Полярні групи амінокислот білків, глікопротеїнів і гліколіпідів також знаходяться на поверхні мембрани. Взагалі ця модель передбачає високий рівень специфічної взаємодії міх компонентами мембрани і відповідно, забезпечує термодинамічний принцип мінімуму вільної енергії системи. Передбачається також асоціація двох або кількох субодиниць інтегральних білків з утворенням специфічних агрегатів всередині мембранию.В таких агрегатах між білками субодиницями можуть утворюватись гідрофільні пори (канали).

Ліпіди всередині мембрани перебувають у "рідкому" стані, що забезпечує їх значну рухливість. У свою чергу ця якість зумовлює динамічність мембрани. Товщина мембрани за "рідинно-мозаїчною" моделлю варіює від товщини білкової області до товщи ни ліпідного бімолекулярного шару. Отже основним структурним елементом мембран є фосфоліпіди, які визначають непроникність мембран для іонів, а отже їх осмотичні і електричні властивості. Обмін клітини речовиною з оточуючим середовищем здійснюють транспортні білки мембран (канали, переносники, насоси).

Холестерин Білок з α-спіраллю занурений в мембрану Гліколіпід Фосфоліпід Глобулярний білок Гідрофобний α-спіральний сегмент білка Олігосахарид

2. Транспорт речовин через мембрани Розрізняють активний і пасивний транспорт нейтральних молекул та іонів через біомембрани. Пасивний транспорт не пов’язаний з використанням хімічної енергії клітини: він здійснюється в результаті дифузії речовин в сторону меншого електрохімічного потенціалу і протікає самовільно. Прийнято розрізняти слідуючі види пасивного транспорту речовин через мембрани: 1) дифузія нейтральних молекул; 2)осмос; 3) фільтрація; 4) дифузія іонів; 5) дифузія через канали (пори); 6) дифузія з допомогою молекул-переносників за рахунок: а) дифузія переносника разом з речовиною в мембрані (рухомий переносник); б)естафетної передачі речовини від однієї молекули-переносника до іншої. Дифузію по механізмах 5 і 6 називають полегшеною дифузією. Поряд з пасивним транспортом в мембранах клітин відбувається переносіонів і молекул в сторону більшого електрохімічного потенціалу. Цей процес, що не являється дифузією і відбувається за рахунок енергії гідролізу АТФ в результаті роботи спеціальних ферментів, які називаються транспортними АТФ-азами – активний транспорт.

- величина, що чисельно дорівнює енергії Гібса одного моля розчиненої речовини, яка знаходиться в електричному полі: Розглянемо механізми пасивного транспорту речовин через біомембрани. Густина потоку речовини при пасивному транспорті підлягає рівнянню Теорелла: (3.1) де u - рухливість часток; С – концентрація. Електрохімічний потенціал (3.2) де F = 96500 Кл/моль – число Фарадея, R – універсальна газова стала, T – температура, φ – потенціал електричного поля; Z – заряд іона електроліту (в одиницях е ), μ0 - хімічний потенціал.
 або (3.4) Рівняння Фіка описує один із найпоширеніших механізмів пасивн...")
(3.3) або (3.4) Рівняння Фіка описує один із найпоширеніших механізмів пасивного транспорту - дифузію.

2.1 Дифузія нейтральних молекул. Дифузія - це процес самостійного повільного проникнення речовини з області з більшою концентрацією в область з меншою концентрацією внаслідок теплового хаотичного руху молекул. Для існування дифузії необхідний градієнт концентрації Закон Фіка для дифузії : (3.5)

Дифузія речовин через ліпідний бішар мембрани викликається градієнтом концентрації в мембрані ( рис.3.4 ). Тоді (3.6) Рис.3.4. де - товщина мембрани. Оскільки виміряти См неможливо, оцінюють См по концентрації цієї речовини в розчині ззовні мембрани С1, або зсередини її С2;
 (3.8) k-коефіцієнт розподілу, показує, яку частину концентрація всереди...")
(3.7) (3.8) k-коефіцієнт розподілу, показує, яку частину концентрація всередині складає від концентрації ззовні. Тоді рівняння Фіка буде: (3.9) (3.10)

2.2 Осмос. Осмос – це дифузія молекул води через напівпроникну мембрану з області меншої концентрації розчиненої речовини в область більшої її концентрації. Сила, яка викликає рух молекул розчинника (води), називається осмотичним тиском. За законом Вант-Гоффа: (3.11) де і – ізотопічний коефіцієнт, який показує, у скільки разів збільшується кількість розчинених часток при дисоціації молекул в електролітах; для неелектролітів і = 1; С – концентрація розчиненої речовини; R – газова стала; T – абсолютна температура розчину.
: (3.12) де - швидкість переносу вод...")
Рівняння осмосу аналогічне рівнянню (3.4): (3.12) де - швидкість переносу води через площу S; р1 і р2 - осмотичні тиски розчинів по обидва боки мембрани; k - коефіцієнт проникності. Осмос відіграє значну роль в біологічних явищах. Наприклад, осмос зумовлює гемоліз еритроцитів у гіпотонічних розчинах. Вода доти проникає всередину клітини, доки гідростатичний тиск всередині клітини не зрівноважить осмотичний тиск. Внаслідок цього клітина набрякає, а мембрана розтягується. Дія деяких послаблюючи лікарських препаратів грунтується на осмосі води в область підвищеної концентрації цього препарату в шлунку

- градієнт тиску в напрямі переносу. Явище фільтрації відіграє важливу роль у переносі води через стінки кровоносних судин, тобто в процесах обміну водою між кров'ю і тканиною. Фільтрація плазми крові в ниркових нефронах приводить до утворення первинної сечі. При порушеннях об міну фільтрація веде до набряків в тканинах. 2.3 Фільтрація. Фільтрація – це рух молекул води через пори у мембрані під дією градієнта статичного тиску. Швидкість фільтрації підлягає закону Пуазейля: ; , (3.13) де u – об’єм фільтрованої води; t - час; x – гідравлічний опір пори радіусом R; h - в’язкість води;

2.4 Дифузія іонів через мембрану На мембрані існує різниця потенціалів, так звані біологічні потенціали, а в самій мембранні - електричне поле. В загальному випадку перерос іонів через мембрану визна чається двома факторами: нерівномірністю їх розпаду і дією електричного поля мембрани, тобто наявністю двох градієнтів - концентраційного і електричного . Підставимо (3.2) в (3.1) і маємо рівняння Нернста-Планка: (3.14) Іншу формулу рівняння Нернста-Планка можна одержати, якщо згадати співвідношення між коефіцієнтом дифузії D і абсолютного температурою: Оскільки можна вважати електричне поле в мембрані постійним , де φм - різниця потенціалів на мембрані, - її товщина. Тоді: (3.15)

. Шляхом математичних перетворень, які ми тут, не приводимо, можна рівнянню (12) надати вигляду: Позначимо (3.16) Вибіркова проникність мембрани залежить від того, яка густина фіксованих зарядів одного знаку в порі. Якщо вона велика, то добре проникають іони протилежного знаку і погано однойменні. Іони H+ і OH- практично в клітину не проникають, оскільки вони адсорбуються поверхнею мембрани, заряджають її і відштовхують інші іони. Проникнення іонів з розчинів підлягає правилу: чим вищий ступень дисоціації, тим менша проникність іонів. Однобічна проникність електролітів та їх нагромадження в клітині зумовлені різницею між вмістом клітини і зовнішнім середовищем.

, і коефіцієнта дифузії речовини в воді D : 2.5 Полегшена дифузія. Дифузія речовини чераз пори (канали) також описується дифузійними рівняннями наявність каналів суттєво збільшує проникність мембрани, ефективний коефіцієнт проникнос ті мембрани в цьому випадку задежить від числа каналів на 1см2 площі мембрани n, радіуса канала r , його довжини (3.17) Ще одне "полегшення" дифузії зв'язане з молекулами-переносниками (Р). Механізм такої дифузії визначається дифузією через мембрану комплексу переносника з речовиною (Ax) в одному напря мі і дифузією вільного переносника в протилежному напрямі. Аx - речовина зовнішнього середовища. При хімічній спорідненості А і Х утворюється комплекс АХ. Схема полегшеної дифузії показана на рис.3.5.

АХ А А А А А А А Х Х Х Х Х Х Х Х Х Х Х Х АХ мембрана Рис3.5.


Рис.3.6. Іонофори є двох видів: переносники і каналоформери. Валіноміцин - переносник. Молекули каналоформерів фіксовані поперек мембрани (рис.3.6). Речовина, яка переноситься, утворює з кож ною фіксованою молекулою комплекс, який розпадається по черзі, поступово, як показано стрілками, молекула А проникає на протилежний бік мембрани.

2.6 Активний транспорт. Тепер можна впевнено сказати, що механізми активного транспорту іонів локалізовані в мембрані, оскіль ки перенесення іонів можна спостерігати на препаратах , в яких немає аксоплазми, але зберігається неушкодженою мембрана. Безсумнівно, що в транспорті іонів беруть участь компоненти мембрани і насамперед фермент Na+, K+ АТФ-аза є основною структурою транспортного механізму, який дістав назву натрій-калієвої помпи і здійснює нагромадження в клітині калію і виведення з клітини натрію.

Рис.3.7 К 3 + Na+, K+- АТФ-аза - це великий мембранний комплекс з складною структурою. Її молекула складається з двох субодиниць - великої, a-ліпопротеїда і меншої, b - глікопротеїда. a - поліпептид виявляє каталітичну активність і може фосфоризуватися в b - глікопротеїд не здатний каталізувати гідроліз АТФ, але виявляє Na -специфічну іонофоретичну активність. Являючись інтегральним мембранним білком Na+, K+ АТФ-аза пронизує мембрану наскрізь і тому частини молекули виступають як в середину клітини ( a одиниця), так і за межі клітини ( b - од.) Активний центр молекули, відповідальний за гідроліз АТФ, повернутий всередину клітини і локалізований на a - поліпептидові(рис.3.7.). присутності АТФ.

L+ Na+ цитоплазма Рис.3.8. Кількісні взаємовідношення в АТФ-азній реакції такі: при розщепленні однієї молекули АТФ в зовнішнє середовище переноситься три іона Na+, а в цитоплазму поступає два іона K+ . АТФ: Na+: K+ = 1:2:3 Отже клітину покидає більше позитивно заряджених частинок, аніж акумулюється в ній. Це створює надлишковий від'ємний заряд всередині клітини і мембранний потенціал на її поверхні. Міжклітинне середовище Хоча конкретні механізми активного транспорту ще точно невідомі, запропоновано багато гіпотез , які пояснюють його. Частина гіпотез грунтується на уявленнях про участь в транспорті спеціальних переносників.

Згідно з однією з таких гіпотез Na+ переноситься з переносником Y. На зовнішній поверхні мембрани переносник Y спонтанно перетворюється в переносник X , здатний зв'язувати тут іони K+ . Калій переноситься на внутрішню поверхню мембрани, вивільняється тут, а переносник X перетворюється в переносник Y. На це перетворення витрачається енергій метаболізму( ).

Сучасні уявлення про механізм дії натрій-калієвої помпи будуються на уявленнях про те, що білки які її утворюють можуть знаходитися в двох різних конформаційних станах. В першій конформації білок зв'язує три іони Na+ тобто виступає в ролі пе реносника Y в другій - два іони K+ ,що відповідає переноснику X . Не треба думати, що в клітині все відбувається саме так. Ця схема одна із спроб наочно подати роботу натрій -калієвої помпи. Ще не встановлено точно природу носіїв проте висловлюються припущення, що комплекс АТФ-аза може виконувати функції справжнього носія. Деякі вчені розглядають функції натрієвої помпи значно ширше. Так, з активним перенесенням може бути пов'язаний транспорт інших речовин. Цей погляд висувають на противагу, традиційному, згідно з яким, скільки речовин транспортується, стільки є транспортних механізмів. Транспорт двох частинок в одному напрямі називають симпортом, а в протилежному напрямку - антипортом.
Схожі презентації











Категорії











