Презентація на тему:
Біохімія печінки. Біотрансформація
Завантажити презентацію










 - найважливіша...")











 і аміноетанолсульфатом (таурином) Кон ...")


 Лужна фосфатаза (ЛФ) (ізоферм...")



 У сироватці крові зростают...")





Біохімія печінки. Біотрансформація
Завантажити презентаціюПрезентація по слайдам:

БІОХІМІЯ ПЕЧІНКИ. Ч. IV Знешкодження токсичних речовин у печінці

Суть реакцій знешкодження токсичних сполук у печінці Внаслідок низки хімічних перетворень (окислення, віновлення, метилювання, ацетилювання, кон югації з рядом сполук) токсичні речовини перетворюються на менш токсичні або нетоксичні завдяки тому, що новоутворені речовини є більш водорозчинними, ніж вихідні, що посилює їх виведення з організму через нирки Є виняткові випадки, коли утворені водорозчинні речовини стають ще більш токсичними або набувають канцерогенних властивостей

Приклади сполук, які знешкоджуються у печінці Чужорідні речовини (ксенобіотики): - харчові додатки - ліки - пестициди - гербіциди - інсектіциди - косметичні засоби - хімічні продукти побутового користування - промислові отрути Ендогенні токсини: - білірубін - аміак - біологічно активні аміни - продукти гниття амінокислот у кишечнику (індол, фенол, скатол) - токсичні гідрофобні жовчні кислоти

2 фази системи біохімічної детоксикації шкідливих речовин у печінці Детоксикація окремих сполук може включати обидві фази або лише одну із них

2 фази системи біохімічної детоксикації шкідливих речовин у печінці: схема

Реакції модифікації Пов язані з функціонуванням монооксигеназних систем гладенького ЕПР Це реакції окислення (гідроксилювання, введення епоксидної групи, утворення сульфоксидів, дезалкілування, дезамінування), відновлення (карбонільної групи, азо- чи нітросполук, дегалогенування), гідролізу ефірів і пептидів (напр., ацетилсаліцилової к-ти), десульфування) В їх результаті у склад неполярних молекул вихідних речовин вводяться функціональні групи -ОН, -СООН, -SH, -NН2, що збільшує їх полярність

Система цитохрому р450. Мікросомальні монооксигенази Після диференційного центрифугування фрагменти мембран ЕПР утворюють т. зв. мікросомну фракцію (або фракцію мікросом), що лишається в рідині над осадом після видалення важких часток, ядер і мітохондрій. В мікросомах печінки локалізований особливий дихальний ланцюг, що містить ферменти окислення - оксигенази. Оксигенази каталізують реакції введення одного (монооксигенази) чи двох (діоксигенази) атомів молекули кисню в субстрати і потребують присутності донорів електронів НАДН, НАДФН та ін. Для мікросомальної системи гідроксилювання печінки характерними є монооксигенази. Монооксигенази каталізують розщеплення сполук різного типу за участю НАДФН та О2; при цьому один атом кисню приєднується до субстрату, а інший звільнюється у складі молекули води: 2НАДФН + R-H + O2 2НАДФ+ +R-ОН +Н2О, або R-H + 1/2O2 R-OH

Компоненти мікросомальної гідроксилюючої системи цитохрому р450 печінки Компонентами ланцюгів мікросомальної гідроксилюючої системи печінки є: ФАД-вмісний флавопротеїн (НАДФН-цитохром р450-редуктаза), цитохром b5 та кінцева монооксигеназа - цитохром Р-450, а також фосфоліпіди й НАДН-цитохром-b5-оксидоредуктаза: НАДФН ФАД цитохром b5 кінцева монооксигеназа (= цитохром Р-450) НАДФН-цитохром р450-редуктаза здійснює транспорт електронів від НАДФН до цитохрому р450, які необхідні для активації комплексу цитохром р450 з субстратом. Це флавопротеїн, що містить одну молекулу ФАД і одну молекулу ФМН, що не пов’язані з білком ковалентно, на один поліпептидний ланцюг. цитохром Р-450 - пояснення у наступних слайдах фосфоліпіди (головним чином фосфатидилхоліни, беруть участь майже в усіх стадіях мікросомального гідроксилювання, підвищують спорідненість субстрату до ферменту, стабілізують активну конформацію цитохрому Р-450). цитохром b5 НАДН-цитохром-b5-оксидоредуктаза

Система цитохрому р450. Типи реакцій, які каталізують мікросомальні монооксигенази Найбільш поширені реакції гідроксилювання по насиченому вуглецевому атому - окислення аліфатичних сполук, гідроксилювання стероїдів, холестеролу, аліциклічних сполук та алкільних бічних ланцюгів. Монооксигенази також каталізують гідроксилювання ароматичних, гетероциклічних сполук, N-гідроксилювання та N-окислення та ін.
 - найважливіша...")
Система цитохрому р450. Цитохром р450 Цитохром р450 (ц/х р450) - найважливіша мікросомальна монооксигеназа, що використовує НАДФН як джерело окисно-відновних еквівалентів В 1958 році Гарфікель та Клингенберг незалежно відкрили гемопротеїн цитохром Р-450; його назва зумовлена тим, що відновлена форма гему зв язує оксид вуглецю (СО) і набуває характерного поглинання світла при 450 нм. Це родина гемопротеїнів з М.м. 50 кД; в різних біологічних системах виявлено понад 300 ізоформ ц/х р450, які різняться за своєю субстратною специфічністю і особливостями первинної структури Цитохром Р-450 бере участь в багатьох стадіях біосинтезу та метаболізму жовчних кислот, стероїдних гормонів, ейкозаноїдів, в утворенні ненасичених жирних кислот Фізіологічне значення існування ізоформ ц/х р450: захист тваринного організму від численних низькомолекулярних ксенобіотиків, що потрапляють у організм. Систему ц/х р450 ще називають “другою імунною системою”, яка є додатковою до основної системи імунного захисту (остання протидіє надходженню в організм чужорідних високомолекулярних сполук біологічного походження) Біосинтез різних ізоформ цитохрому р450 кодується кількома підродинами генів, які у ссавців позначаються як CYP (від англ. cytochrome p450). В геномі людини за синтез всіх ізоформ ц/х р450 відповідають понад 100 генів (CYP1, CYP2, CYP3,...). Клінічний інтерес мають ізоформи ц/х р450, причетні до біотрансформації численних лікарських препаратів, зокрема, CYP1А2, CYP2С8 - 10, CYP2С-19, CYP2D6, CYP2Е, CYP3А4 Окислена форма Р-450 відновлюється і активує кисень. Цитохром Р-450 виконує подвійну функцію - 1) зв’язує субстрат гідроксилювання, 2) на ньому відбувається активація кисню

Схема функціонування мікросомальної гідроксилюючої системи (за R. Estabrook et al) І стадія: взаємодія субстрату з окисленою формою ц/х р450; утворення фермент-субстратної форми при цьому супрводжується переходом гемового заліза із низько- у високоспиновий стан ІІ стадія: відновлення ферментно-субстратного комплексу першим електроном, який надходить від НАДФН за участю флавопротеїну - НАДФН - цитохром Р-450 - редуктази. Передбачають у цьому участь цитохрому в5, що міститься в мембранах ендоплазматичного ретикулуму. ІІІ стадія: утворення потрійного комплексу SH-Fe2+-О2. Кисень приєднується до відновленого фермент-субстратного комплексу з великою швидкістю

Схема функціонування мікросомальної гідроксилюючої системи (за R. Estabrook et al) (продовження) Іv стадія: потрійний комплекс відновлюється другим електроном, який передається з НАД-специфічного ланцюгу переноса, що містить НАДН - цитохром b5 - редуктазу і цитохром b5. Є дані про взаємодію чи спряження двох ланцюгів переноса електронів - НАДФН- та НАДН-специфічного - через цитохром b5. V стадія - складні внутрішньомолекулярні перетворення відновленого потрійного комплексу та його розпад із звільненням гідроксильованого похідного та води

Регуляція активності ц/х р450

Оцінка стану мікросомальної системи Гексаналова проба: у тварин викликають “гексаналовий сон” і реєструють тривалість сну. Зменшення її свідчить про індукцію ферментів, пролонгація - про інгібіцію Антипіринова проба - грунтується на визначенні у сечі вмісту метаболітів антипірину - 4-аміноантипірину й n-ацетил-4-аміноантипірину, які взаємодіють із фенолом і утворюють сполуки типу індофенолу. Посилення метаболізму антипірину свідчить про індукцію монооксигеназної системи, а гальмування - про її пригнічення Захворювання печінки супроводжуються зниженням детоксикаційної функції, що може бути причиною підвищення чутливості до ряду ліків, які біотрансформуються системою ц/х р450

Реакції кон югації Кон югації підлягає більшість ксенобіотиків, що мають функціональні групи, які утворилися в стадії модифікації Кон югація - це енергозалежний процес Значна частина цих реакцій відбувається на мембранах ЕПР, проте деякі з них протікають в цитоплазмі і в інших структурах клітини Найпоширеніші реакції кон югації: - глюкуронідна - сульфатна - кон югація з глутатіоном - кон югація з амінокислотами (глутаміном, гліцином, тощо) - метилювання і ацетилювання Із печінки кон югати виводяться рецептор-залежною екскрецією в жовчні капіляри або через нирки

Кон югація з глюкуроновою кислотою Активною формою глюкуронової кислоти є уридиндифосфатглюкуронова к-та (УДФГК), що утворюється при окисленні УДФ-глюкози НАД-залежною УДФГ-дегідрогеназою: УДФГ +2НАД УДФГК +2НАДН2 Загальна реакція глюкуронідної кон югації: R-OH + УДФГК R-О-глюкуронід + УДФ (фермент - УДФ-глюкуронілтрансфераза) В залежності від хімічної природи субстрату - О-, N-, S-глюкуронування: R-NH2 +УДФГК R-NH-глюкуронід +УДФ R-SH + УДФГК R-S- глюкуронід +УДФ Характерна для чужорідних сполук, спостерігається у білірубіну, тироксину і естрадіолу, продуктів гниття білків у кишечнику

Приклад реакції кон югації з глюкуроновою к-тою

Кон югація з сірчаною кислотою Активна форма сірчаної кислоти, що використовується в реакціях кон югації - 3 -фосфоаденозин-5 -фосфосульфат (ФАФС) Ферменти, що здійснюють ці реакції - сульфотрансферази Загальна реакція сульфатної кон югації: R-OH + ФАФС R-OSO3H +ФАФ Сульфатній кон югації підлягають стероїдні гормони і продукти їх метаболізму, продукти гниття білків у кишечнику (фенол, скатоксил, індоксил, крезол), сторонні речовини

Кон югація з глутатіоном Глутатіон - це трипептид гама-глутамініл-цистеїніл-гліцин В основі - здатності глутатіону вступати до реакції кон югації зі сполуками, які містять електрофільні атоми вуглецю, здатні реагувати з -SH-групами за участю глутатіон-S-трансфераз (ГТаза) Механізм. Спочатку ГТази каталізують взаємодію субстрату (R-X, де Х = -ОН чи ін. група) з відновленим глутатіоном (глутатіон-SH): R-X +глутатіон-SH R-S-глутатіон +ХН (R-S-цистеїн-гліцин-глутамат) Від глутатіонового кон югату далі відокремлюються послідовно глутамінова к-та і гліцин з утворенням кон югатів з цистеїном, які виводяться через нирки або перетворюються у меркаптурові кислоти, які теж виводяться із сечею Цим шляхом знешкоджуються сторонні речовини, різні за структурою, а також - у невеликій к-ті - білірубін, естрадіол, простагландини і лейкотрієни

Кон югація шляхом ацетилювання Таким чином знешкоджуються сторонні ароматичні аміни, ароматичні амінокислоти, сульфаніламідні продукти Загальна схема: R-NH2 +CH3-CO-SKoA R-NH-CO-CH3 +KoA-SH ацетил-КоА

Кон югація шляхом метилювання Цей тип кон югації характерний для амінів, фенолів, тіолових сполук, неорганічних сполук сірки, селену, ртуті, срібла, катехоламінів (адреналін, норадреналін), вітаміну РР Донором метильної групи виступає S-аденозилметіонін Ферменти реакції - метилтрансферази Загальна реакція: R-ОН +S-аденозилметіонін R-O-CH3 +S-аденозилгомоцистеїн
 і аміноетанолсульфатом (таурином) Кон ...")
Кон югація з амінокислотами (гліцином) і аміноетанолсульфатом (таурином) Кон югація з гліцином характерна для ароматичних і гетероциклічних карбонових кислот, здійснюється у 2 стадії: І стадія: утворення КоА-похідного сторонньої карбонової кислоти (напр., бензойної): ІІ стадія: пептидна кон югація з амінокислотою гліцином: Кон югація з таурином СООН +АТФ +КоА-SH СО-SКоА +АМФ +ФФн СО-SКоА + Н2N-СН2-СООН CO-NH-СН2-СООН +КоА-SH

Найважливіші реакції кон югації, що застосовуються для оцінки стану детоксикаційної функції печінки Проба Квіка-Пителя: після навантаження бензоатом натрію у сечі визначають вміст утвореної гіпурової кислоти. Введений в організм бензоат натрію взаємодіє з ендогенним гліцином з утворенням гіпурової кислоти: При паренхіматозних ураженнях печінки цей показник знижується Визначення екскреції тваринного індикану - сполуки, що утворюється в печінці на основі індолу. Індол є похідним L-триптофану і синтезується в товстому кишечнику під впливом ферментів мікроорганізмів в процесі гниття білків у кишечнику. Утворення тваринного індикану шляхом реакції сульфатної кон югації відбувається за схемою: L-триптофан індол індоксил індоксилсульфат тваринний індикан (це К-сіль індоксилсульфату)

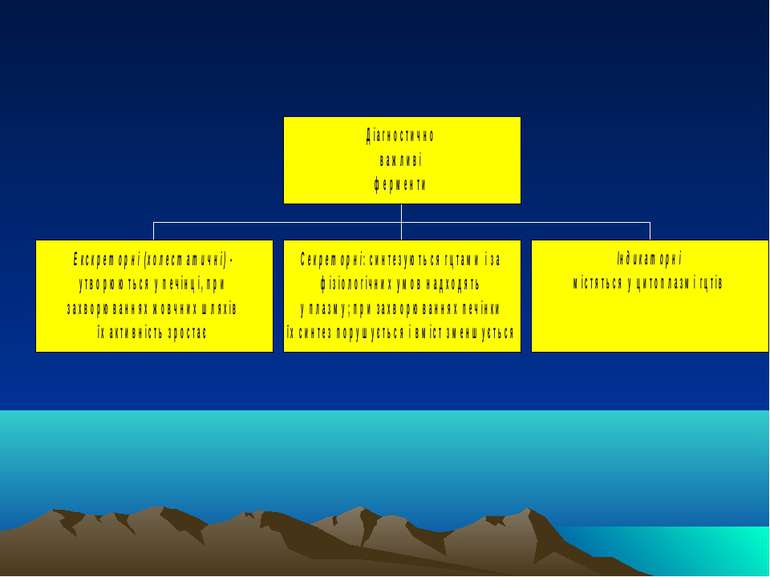
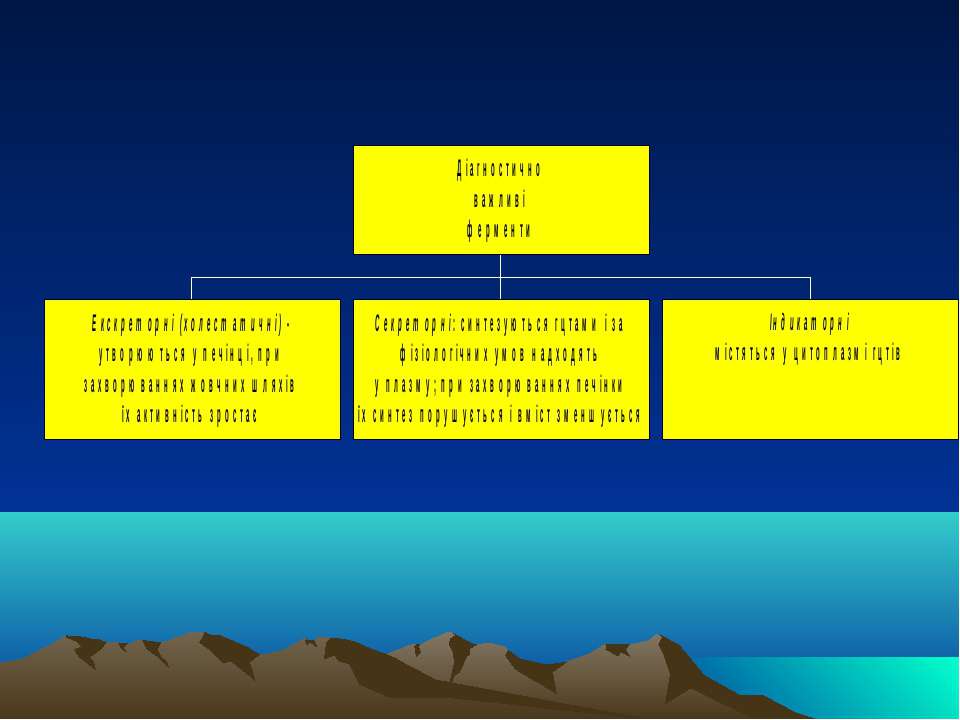
Біохімія печінки. ч.V. Сироваткові ферменти, важливі для діагностики захворювань печінки. Дослідження функціонального стану печінки за синдромним принципом
 Лужна фосфатаза (ЛФ) (ізоферм...")
Екскреторні ферменти Лейцинамінопептидаза (ЛАП) Лужна фосфатаза (ЛФ) (ізофермент, специфічний для печінки; інші ізоферменти - плацентарний, кістковий, кишечний, нирковий). Каталізує гідроліз складноефірних зв язків у моноефірах фосфорної к-ти і органічних сполук. Різке збільшення активності спостерігається за умов механічної жовтяниці, менщ виражене - при гепатиті і цирозі; часом підвищення активності ЛФ є майже єдиною ознакою злоякісних новоутворень печінки Гама-глутамілтранспептидаза (= гама-глутамілтрансфераза) - каталізує перенос гама-глутамілового залишку з гама-глутамілпептиду на акцепторний пептид чи L-амінокислоту. Збільшення її активності завжди пов язане з ушкодженнями печінки: при холестичних станах (паралельно із ЛФ), при прийомі деяких антисудомних ікарських препаратів та алкоголю, за алкогольного гепатиту (застосовують як тест на хронічний алкоголізм) .

Секреторні ферменти Прокоагулянти (зокрема, фібриноген, протромбін, проакцелерин і проконвертин, антигемофілічні фактори) - тому одним із найцінніших прогностичних показників є визначення протромбінового часу (це проба на зсідання крові), а також вміст прокоагулянтів у плазмі крові Холінестераза печінки (ін. назва - псевдохолінестераза, ХЕ) - здійснює розщеплення ефірів холіну (ацетилхоліну, бензоїлхоліну, сукцинілхоліну тощо) за схемою (для ацетилхоліну): СН3СООСН2СН2N+(СН3)3 + Н2О СН3СООН + НОСН2СН2N+(СН3)3 ацетилхолін оцтова к-та холін Зниження активності ХЕ в сироватці крові відображує тяжкість і поширеність клітин печінки за гострого гепатиту, цирозу, онкологічних захв-х печінки. Показник важливий для спостереження за протіканням хвороби і для прогнозу хронічних захворювань печінки Церулоплазмін (мідна оксидаза) - основна оксидаза плазми крові, яка належить до групи білків гострої фази. Вона здійснює транспорт міді для синтезу супероксиддисмутази і цитохромоксидази; регулює рівень біогенних амінів, у т.ч. катехоламінів; стимулює фагоцитарну і бактеріцидну активності нейтрофілів і моноцитів; є ферроксидазою - окислює Fe++ до Fe+++, який здатний вбудовуватися у трансферин і в такий спосіб транспортуватися у кістковий мозок. Зменшення його вмісту - при недостачі синтезу печінкою (некроз печінки, хвороба Вільсона або гепатолентикулярна дегенерація). Рівень ЦП значно зростає за різних інфекційних захворювань (це білок гострої фази), у т.ч. за гепатитах.

Індикаторні ферменти Трансамінази: АлАТ і АсАТ. В нормі співвідношення АсАТ до АлАт (коефіцієнт де Рітіса) = 1,3 - 1,4. При жовтяничних та безжовтяничних формах гострого гепатиту, некрозі печінки, лікарських її ураженнях, за вірусного гепатиту, метастазах у печінці, гепатомах, інфекційного мононуклеозу, при цирозах печінки і хрон. гепатитах в періоди загострень збільшується активність трансаміназ, особливо АлАТ (тобто коефіцієнт де Рітіса стає нижчим за норму). За алкогольного гепатиту виразніше зростає АсАТ. Максимальна активність цих ферментів - при переході хрон. гепатиту в цироз. Альдолаза (фруктозо-1,6-дифосфатальдолаза) - гліколітичний фермент, що міститься у скелетних м язах і печінці та каталізує обернену реакцію: фруктозо-1,6-дифосфат діоксиацетонфосфат + гліцеральдегід-3-фосфат При ураженні зазначених органів з являється у великій кількості у крові. Її активність зростає у преджовтяничному періоді хвороби Боткіна через активацію вірусом перших етапів процесу гліколізу. Фруктозо-1-фосфатальдолаза - каталізує наступну реакцію: фруктозо-1-фосфат діоксиацетонфосфат + гліцеральдегід Визначення активності фермента служить специфічним показником ушкодження паренхіми печінки. В десятки разів зростає при гострому гепатиті, в 2-3 рази - за токсичного ураження печінки. При новоутвореннях і захворюваннях жовчовивідних шляхів цей показник не змінюється ЛДГ - гліколітичний фермент, що каталізує реакцію перетворення лактату на піруват: лактат + НАД піруват + НАДН Є 5 ізоформ цього ферменту, що відрізняються за субодиничним складом (М-с/о і Н-с/о, від muscle і heart): ЛДГ1 (Н4) і ЛДГ2 (Н3М) - в серцевому м язі, ЛДГ3 (Н2М2) - в легенях, селезінці лімфовузлах, ЛДГ4 (НМ3) і ЛДГ5 (М4) - в печінці і скелетних м язах. За гострого гепатиту різко зростає активність печінкових форм (в 6-8 разів) і зменшується - двох перших. ЛДГ5 зростає при цирозі, всіх типах гепатиту, пасивному застої у печінці. В нормі у сироватці: ЛДГ1 -31%; ЛДГ2 - 40%; ЛДГ3 - 17%; ЛДГ4 - 8%; ЛДГ5 - 4%

Дослідження функціонального стану печінки за синдромним принципом На основі патоморфологічного принципу розрізняють низку синдромів, пов язаних зі змінами активностей ряду ферментів: - синдром цитолізу - синдром холестазу - синдром гепатоцелюлярної недостатності - мезенхімально-запальний синдром - синдром регенерації і пухлинного росту - синдром шунтування печінки
 У сироватці крові зростают...")
Синдром цитолізу (порушення цілісності гепатоциту) У сироватці крові зростають активності: - АсАТ і АлАТ - ЛДГ - фруктозо-1-фосфатальдолази - гамаглутамілтранспептидази - орнітилкарбамоїлтрансферази - сорбітолдегідрогенази - глутаматдегідрогенази

Синдром холестазу в сироватці крові зростають активності: - ЛАП; - гама-глутамілтранспептидази - ЛФ - КФ (кислої фосфатази) - 5 -нуклеотидази Також зростає вміст ЖК, білірубіну (загального за рахунок кон югованого), холестеролу крові

Синдром гепатоцелюлярної недостатності Одна з основних ознак - зниження у сироватці крові активності холінестерази. Іншими індикаторами є: - бромсульфалеїнова і вофавердинова проби - антипіринова і кофеїнова проби - проба Квіка-Пителя - тести на прокоагулянти (фібриноген, протромбіновий індекс, проакцелерин і проконвертин, антигемофілічні фактори), вільний гепарин - дослідження вмісту альбумінів сироватки крові, альфа-1-антитрипсину, церулоплазміну

Індикатори мезенхімально-запального синдрому Синдром пов язаний із пошкодженням мезенхіми і строми печінки (фіброзна оболонка +портальний тракт +внутрішньопечінкова сполучна тканина) Основні індикатори: - гама-глобулін сироватки крові - осадові реакції (тимолова і сулемова проби) - імуноглобуліни A, G, M, D сироватки крові - вміст продуктів обміну сполучної тканини (оксипролін плазми крові й сечі, проколаген-ІІІ-пептид (=проколаген печінки))

Індикатори синдрому регенерації і пухлинного росту При пухлинах, зокрема, при гепатоцелюлярній карциномі, у сироватці крові з являється альфа-фетопротеїн

Індикатори синдрому шунтування печінки До них належать речовини, які в нормі головним чином надходять із кишечника у систему ворітної вени і в печінку: - аміак та його похідні - феноли - амінокислоти тирозин, фенілаланін, триптофан, метіонін - жирні кислоти з коротким ланцюгом В патологічних умовах, при розвитку венозних колатералей, ці сполуки потрапляють у систему загального кровотоку, не надходячи до печінки, і стають показником викиду венозної портальної крові. Шунтування печінки таким чином спричиняє кишкову інтоксикацію.
Схожі презентації











Категорії











